Progress in the miniaturization of CRISPR-Cas systems
CRISPR-Cas系统的小型化研究进展

摘要
CRISPR-Cas基因编辑技术由于其简便性和高效性,已被广泛应用于生物学、医学、农学等领域的基础与应用研究。目前广泛使用的Cas核酸酶均具有较大的分子量(通常大于1000个氨基酸),而广泛应用于基因治疗中的腺相关病毒(AAV)载体的承载容量却十分有限,在容纳CRISPR核酸酶与gRNA的编码序列之余往往难以承载更多其他功能元件,如碱基编辑、转录调控、多基因编辑等相应元件,这严重限制了其在基因治疗等领域的应用。使用紧凑型Cas蛋白变体的CRISPR-Cas系统可能有助于用AAV产生和传递基因组编辑和调节工具到人类细胞。因此,小型化的CRISPR-Cas系统开发是解决这一技术难题的重要途径,本文主要概括了基于Cas9、Cas12和Cas13蛋白系统在小型化方面的研究进展,包括筛选新型Cas蛋白、缩减蛋白结构域以及引导RNA的改造等,旨在为开发微型精准基因编辑和调控工具提供新思路。目前小型化的CRISPR-Cas系统的局限性主要体现在蛋白分子量的大小和基因编辑的效率、特异性不可兼得上,在未来的研究中若能解决这一问题,获得更小型化的结构域,相信不仅能够优化该系统在体内的传递,更有望为临床带来高效率且低损害的治疗方法。
CRISPR-Cas系统是微生物抵抗移动遗传元件(mobile genetic elements,MGE)入侵的关键适应性免疫成分,也是当前尖端基因组工程技术的核心[1]。该系统设计简单,可编辑位点广泛,目前已成功在动植物基因组中进行编辑,包括使用多效应蛋白的1类系统和使用单蛋白效应域的2类系统。由于单蛋白效应域的优势,2类系统是用于生物学研究和翻译应用的最广泛的CRISPR工具,其可进一步细分为Ⅱ、Ⅴ和Ⅵ三种类型[2]。在2类系统的Cas蛋白中,多数Ⅱ型Cas9和Ⅴ型Cas12具有RNA引导的DNA内切核酸酶活性,而Ⅵ型Cas13显示出RNA靶向和切割活性,Cas9、Cas12和Cas13主要出现的靶向切割特点已展示于图1。目前三种工具因切割特性已分别被应用于不同的领域,例如小型Cas9和Cas12常应用于基因治疗及基因组编辑;Cas13的某些变种已经被作用于RNA传感器,用于检测特定的RNA序列,并且已成功应用于各种病原体的检测[3],这三种不同Cas蛋白的特点已总结于表1。
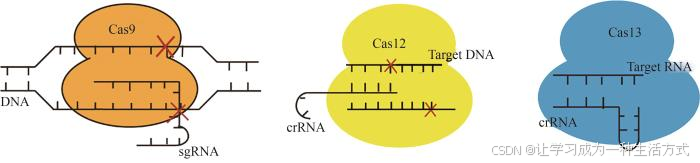
图1 Cas9、Cas12和Cas13的靶向切割特点
Table 1 Comparison of the characteristics of Cas9, Cas12 and Cas13
| 特点 | Cas9 | Cas12 | Cas13 |
|---|---|---|---|
| 靶标类型 | DNA | DNA | RNA |
| 切割模式 | 产生平末端的DNA双链断裂 | 产生黏性末端的DNA双链断裂(部分产生单链DNA切割断裂) | 产生单链RNA断裂 |
| PAM序列(用于识别并结合目标) | 通常需要一个特定的PAM序列 | 需要PAM序列(不同于Cas9) | 不需要PAM序列,直接靶向于RNA |
| 优势 | 广泛应用,工具资源丰富,操作简单 | 因其需要更小的PAM及其黏性末端而更容易进行编辑 | 针对RNA的编辑能力更强 |
| 适用场景 | 基因编辑及调控、染色质成像;DNA剪切、修复和替换;转录调控[4] | 基因编辑及调控、染色质成像;特异性即时检测[5] | RNA编辑、调控翻译水平、RNA修饰;基因沉默;抑制病毒RNA复制[6] |
| 实际应用案例 | 输血依赖性β地中海贫血(TDT)和镰状细胞病(SCD)[7] | 检测COVID-19[8]、SARS-CoV-2 | 开发SHERLOCK技术,检测寨卡和登革热病毒的特定菌株[9] |
目前已有大量实验证实,CRISPR-Cas系统衍生工具在转录调控、染色质成像、碱基编辑、疾病诊断、基因功能筛选、疾病模型建立、免疫治疗、基因治疗以及活细胞标记和成像方面具有应用潜力[10]。
AAV载体由于其低致病风险、低免疫原性和广泛的组织趋向性,在临床疾病治疗中得到广泛应用,是CRISPR-Cas系统重要的递送载体[11-13]。然而,AAV负载能力有限,常用的Cas蛋白的大小限制了系统在治疗应用中的效用,影响基因编辑的效率和特异性[14]。其他的递送方法包括显微注射[15]、电穿孔[16]和水动力尾静脉注射[17]等,递送效率也会受递送蛋白分子量的影响。
Cas蛋白实现功能需多个阶段,包括核糖核蛋白(RNP)的形成、目标核酸的发现、DNA解旋的启动、gRNA和目标序列之间的互补碱基配对;转录过程中R环(一条DNA:RNA杂交链和一条被置换出的单链DNA组成的三链核酸结构)的形成以及最终对目标核酸的切割[18]。这些广泛的功能需要多个不同的“功能域”来承担[19]。目前已知,对Cas蛋白的小型化研究包括结构域的缩减和筛选新型蛋白。其中,结构域缩减是相对较简单且有效的方法,通过删除或缩减效应蛋白中较大的结构域,保留其功效所需要的最小结构域,使小型化蛋白可以在维持其基因编辑能力的同时,尽可能地减少所协同的不良反应;此外,还可通过缩减gRNA序列来实现小型化。本文将主要从筛选新型蛋白、缩减结构域、缩减gRNA序列这三个方面总结对CRISPR系统进行小型化研究的进展。
1 Cas9的小型化研究
Cas9是Ⅱ型系统的标志性蛋白,与经过加工的crRNA和反式激活的crRNA(tracrRNA)组成的双引导RNA结构结合。Cas9利用crRNA 5'端约20个核苷酸片段结合靶向DNA序列,包括核酸酶结构域HNH和RuvC分别切割靶标dsDNA的互补和非互补DNA链[20-21],以及识别结构域REC、PAM相互作用结构域PI等[22](图2)。对Cas9更多小型化研究总结于表2。
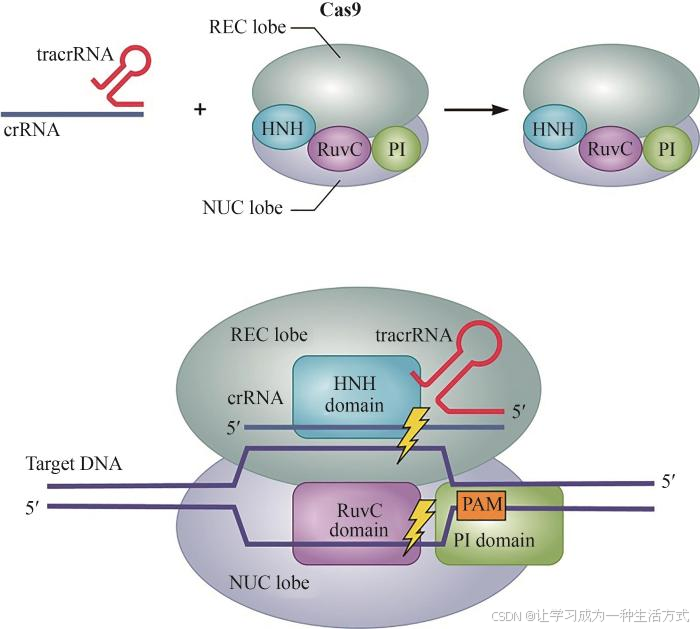
图2 Cas9结构示意图
Table 2 Miniaturization studies of Cas9
| 蛋白 | 分子量/kDa | PAM | spacer/nt | tracrRNA | 直接重复 序列(DR) | 切割dsDNA | 切割ssDNA | 实验细胞/细菌 |
|---|---|---|---|---|---|---|---|---|
| SpCas9[24] | 160 | 5′-NGG-3′ | 42 | Y | 36 | Y | N | HEK293T |
| Nme1Cas9[25] | 162.4 | 5'-N4GAYW/N4GYTT-3'或5'-N4GTCT-3' | 24 | Y | 24 | Y | N | Neisseria meningitidis |
| SaCas9[26-27] | 126 | 5′-NNGRRT-3′ | 21~23 | Y | 19-24 | Y | N | Staphylococcus aureus |
| CjCas9[28] | 108 | 5′-N4RYAC-3′ | 22 | Y | 35 | Y | N | Campylobacter jejuni |
| mini-SaCas9[29] | 100 | 5′-NNGRRT-3′ | 20 | N | 24 | N | N | Staphylococcus aureus |
| Nme2Cas9[30] | 160 | 5′-N4CC-3′ | 22~24 | Y | 24 | Y | N | Neisseria meningitidis |
| SauriCas9[31] | 118 | 5′-NNGG-3′ | 20 | Y | 36 | Y | N | Staphylococcus auricularis |
| BlatCas9[32] | 120 | 5′-N4CNAA-3′ | 17~24 | Y | 24 | Y | N | HEK9T |
| MISER Cas9[33] | 100 | 5′-NNRGAA-3′ | 42 | Y | 36 | N | N | Streptococcus pyogenes |
| SchCas9[34] | 115 | 5′-NNGR-3′ | 21 | Y | 32 | Y | N | — |
| Nsp2Cas9[35] | 117 | 5′-N4C-3′ | 22~26 | N | 23 | Y | N | — |
| IscB[36-37] | 54 | 5′-NWRRNA-3′ | 20 | Y | 206 | Y | N | — |
注:Y—C+T;N—A+C+G+T
1.1 筛选新型Cas9亚蛋白
Ran等[26]对由Cas9酶和嵌合sgRNA(由crRNA和tracrRNA衍生)两个组件组合就能进行真核生物基因组编辑的CRISPR-Cas系统进行分析,将Cas9的600多个同源物按照酶的特征蛋白大小1350 aa和1000 aa分为两个长度组,从中分别选取出能够显著截断REC结构域的6个较小的Cas9同源物,发现这6个同源物中的其中一个——SaCas9可以编辑基因组,其效率与SpCas9相似,但长度较SpCas9短1 kb,仅含有19~22 nt的DR序列和20 nt的间隔序列(spacer)。将SaCas9及其sgRNA表达盒包装到一个AAV载体中,成功针对在小鼠肝脏中的调节胆固醇基因——Pcsk9实现了>40%的基因修饰,此项实验证明了SaCas9介导的体内基因组编辑可能是特异且高效的。Hu等[31]在2018年筛选出体积为1061 aa的SauriCas9蛋白,该蛋白PAM序列为NNGG,团队将SauriCas9与SaCas9进行平行比较,发现两者切割效率相近。同年,该团队还发现BlatCas9(1092 aa)蛋白能够在哺乳动物细胞内产生切割酶活性,其PAM识别序列为N4CNDD[32],团队在后续实验中证实,在条件相同的情况下,SpCas9出现了12个脱靶位点,而BlatCas9只出现了1个。2021年,王永明团队[34]鉴定出新型SchCas9蛋白(1054 aa),该蛋白识别范围较广,可识别以NNGR为PAM的序列。2022年,该团队从Nme1Cas9家族相近的29种蛋白中鉴定出25种能够在细胞内发挥功能的小型蛋白[35],其中Nsp2Cas9蛋白识别的PAM序列是N4C,其蛋白切割效率与Nme1Cas9相近,脱靶率低,团队在三个基因位点上对其进行脱靶风险评估,发现Nsp2Cas9只在一个位点上表现出小幅脱靶。除上述提到的通过表达和纯化以及体外生物信息学分析筛选亚蛋白的方法外,还能够通过体外DNA切割实验、结构生物学研究以及活体模型验证等来筛选Cas9亚蛋白。这些亚蛋白的识别为小型Cas9工具箱的拓展开辟新的路径,并且其中一些亚蛋白有望成为具有巨大潜力的基因编辑工具。
1.2 蛋白质工程改造
Altae-Tran等[38]证明Isc家族是Cas9蛋白的远古祖先,拥有更小的尺寸以及更广泛的生物界分布,并且已经在哺乳动物细胞中证明该家族蛋白有Cas9类似的基因编辑功能。通过高分辨率结构解析,Schuler等[36]发现IscB的大小约为Cas9的2/5,IscB与1个约200~400 nt的非编码RNA(ωRNA)结合,从而实现对特定DNA序列的识别靶向和切割,ωRNA发挥着crRNA和tracrRNA的综合作用,指导dsDNA的裂解,其PAM序列也被称为TAM(target adjacent motif)。通过用ωRNA替换较大Cas9中的蛋白质成分,IscB蛋白核心化学反应(DNA切割)中心仍然存在。研究人员进一步在不影响IscB活性的情况下删掉了55 aa,使最后的蛋白仅有450 aa,这是目前所知的最小Cas9同源体,且其对人类染色体中DNA并未产生较大影响。这项研究说明,通过使用人工合成的RNA和IscB蛋白,可以使Cas9蛋白组件更加简单且易于优化,从而为CRISPR-Cas9技术的应用提供了更多的选择,同时还能更好地控制CRISPR-Cas9系统的功效和安全性。
为了获得更小的Cas蛋白,还可以通过缩减结构域,即在保持其基因编辑的效率和特异性的同时减小Cas9的大小。Shams等[33]通过迭代凝胶排阻的方法最小化技术(minimization by iterative size-exclusion recombination,MISER),用于全面地对一个蛋白质进行最大程度的删减,这一删减使CRISPR蛋白Cas9生成了一个缺失现象。研究发现,dSpCas9可以在REC2、REC3、HNH和RuvC结构域的大的单次缺失中不受影响,正常地在体外和体内发挥作用,而且这些缺失可以堆叠在一起,能设计出最小的、与DNA结合但不切割的效应蛋白MISER Cas9,该蛋白仅包含880 aa,约为原始SpCas9大小的2/3。Ma等[29]根据结构信息,通过删除野生型dSaCas9保守的HNH和/或REC-C结构域,设计了一组缩短0.5 kb的紧凑的Cas9衍生物,该衍生物保留了高效的DNA结合活性。为了增强基因组编辑的准确性,该团队在dCas9的N端与FokⅠ核酸酶结构域在PAM方向上进行了融合。在双gRNA的作用下,该结构结合并执行DNA切割。然而,当spacer的距离或目标内源基因发生变化时,每个FokⅠ融合Cas9的DNA切割效率会有所差异。因此,还需要更多研究来测定这种效率差异,并为不同情境下的DNA切割效率寻找最佳方案。以上实验表明,通过对Cas9蛋白的改造和优化,研究人员可以进一步提高基因编辑的特异性和效率,从而扩大该技术在基因治疗、基因工程和基因研究等领域的应用范围。然而这些技术仍处于研究阶段,需要更多的研究来评估和优化它们在不同情境下的DNA切割效率和稳定性,并确保这些技术在实际应用中的安全性和有效性。
1.3 gRNA改造
在自然系统中,Cas9切割DNA需要两条RNA,包括5′端含有与靶标DNA序列配对的CRISPR RNA(crRNA)[39],以及一条独立的反式激活crRNA(trans-activating crRNA,tracrRNA),其5′端与crRNA的3′端序列反向互补,3′端含有内部的发卡结构。2012年,Jinek等[24]最先对Cas9的DNA切割活性进行了全面的研究,研究人员选择了SpCas9的tracrRNA∶crRNA系统,把crRNA和tracrRNA直接连成一条长约120 nt的RNA,并使用体外Cas9切割系统,通过一系列序列截断,分析了tracrRNA∶crRNA中对Cas9切割活性最重要的区域,研究发现crRNA仅保留1~32 nt、tracrRNA保留23~48 nt的嵌合体RNA(58 nt)可以有效完成Cas9切割。同样,Mali等[40]直接使用了chimericRNA进行真核细胞内的基因编辑实验,其中crRNA保留了1~32 nt、tracrRNA保留了23~79 nt(89 nt),该实验亦有效完成了目的基因的切割。以上实验通过对核酸序列的缩减实现了gRNA成熟的新途径,达到了优化Cas9蛋白大小的目的。
在基础和应用研究中,经常需要对多个基因组基因座进行设计,专门优化的gRNA能够更加配合小型化Cas9的需求,提高基因编辑效率。然而,其多重应用仍然受到其gRNA处理效率和承载量的限制。通常,有两种方法可用于表达多种gRNA:一种方法是用单个RNA聚合酶启动子转录每个gRNA;另一种方法是使用一个启动子在一个转录物中转录所有gRNA,然后通过不同的策略处理以释放单个gRNA。为了进一步减小转基因大小,Zhang等[41]和Mefferd等[42]先后使用约70 nt的tRNAGly启动子取代约2500 nt的RNA聚合酶Ⅲ启动子,用来表达高水平的tRNA∶sgRNA融合转录物,这些转录物被内源性tRNase Z有效而精确地裂解,释放出sgRNA。这一新的sgRNA通过对序列的缩减表达策略极大地促进有效的基于AAV的Cas9/sgRNA载体构建,避免了载体大小的限制。在针对靶向基因特异性设计gRNA的同时,还能减少Cas9蛋白的大小,这为基因编辑的进一步广泛应用提供可能性。
2 Cas12的小型化研究
与Cas9相反,Cas12家族中的Cas12a具有切割pre-crRNA的能力[43],可用于多基因编辑[44]。Cas12a是由识别叶(REC)和核酸酶叶(NUC)组成的双叶分子结构[45-46],同Cas9相比较,它缺乏Cas9靶链切割所需的核酸酶结构域HNH[44],故而由RuvC结构域同时裂解dsDNA的目标链和非目标链,并在PAM位点下游产生交错的末端,这有助于精确插入,利于更有效、准确地整合DNA片段[44](图3)。表3总结了Cas12家族系统中部分小型化蛋白特点。
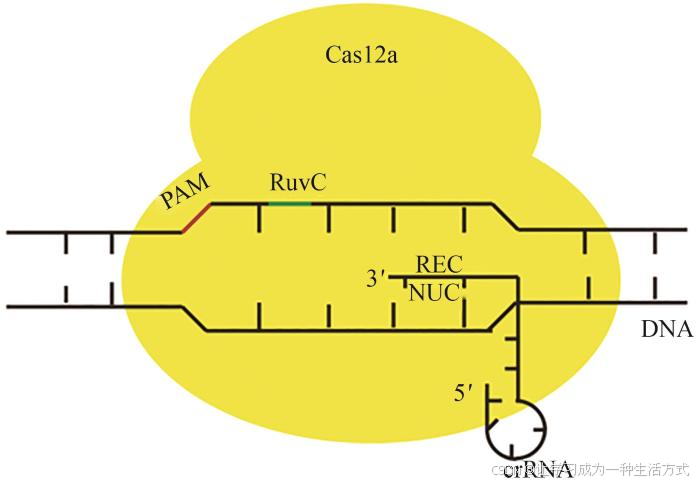
图3 Cas12a结构示意图
Table 3 Miniaturization studies of Cas12
| 蛋白 | 分子量/kDa | PAM | spacer/nt | tracrRNA | 直接重复 序列(DR) | 切割dsDNA | 切割ssDNA | 实验细胞/细菌 |
|---|---|---|---|---|---|---|---|---|
| Cas12d(CasY)[47] | 132 | 5′-TA-3′ | 17 | Y | 26 | Y | — | uncultivated microbes |
| Cas12e5(CasX)[47-48] | 108 | 5′-TTCN-3′ | 20 | Y | 23 | Y | — | uncultivated microbes |
| FnCpf1(Cas12a)[49] | 147 | 5′-KYTV-3′ | 20 | N | 14 | Y | Y | — |
| AaCas12b(C2c1)[50] | 125 | 5′-TTN-3′ | 20 | Y | 37 | Y | Y | Alicyclobacillus acidiphilus |
| Cas12c(C2c3)[51-52] | 133.09~146.3 | 5′-TG-3′或5′- TN-3′ | 17-18 | Y | 17 | Y | Y | HEK293T |
| Cas12g[51] | 79.2~91.3 | 5′-NA-3′ | 30 | Y | 36 | N | Y | HEK293T |
| Cas12h[51] | 95.7~101.64 | 5′- RTR-3′ | — | N | — | Y | Y | HEK293T |
| Cas12i[51] | 113~120 | 5′ TTN-3′ | 28 | N | 24 | Y | Y | HEK293T |
| Cas12j[53-54] | 70~80 | 5′-TBN-3′或5′-TTN-3′ | 14-20 | N | 26 | Y | Y | HEK293T |
| CasMINI[55] | 58 | 5′-TTTR-3′ | 23 | Y | 26 | Y | Y | TRE3G-GFP HEK293T |
| UnCas12f[56-57] | 44~77 | 5′-TTTA-3′ | 34-39 | Y | 37 | Y | Y | — |
注:Y—C+T;N—A+C+G+T
2.1 筛选新型Cas12亚蛋白
Yan等[51]更深入地研究整个Cas12家族,以寻找具有新功能的效应器,建立并搜索了一个包含近30万个假定的Ⅴ型系统的CRISPR-Cas元素的数据库——只有一个预测的RuvC核酸酶域的核酸酶。该团队在大肠杆菌中使用了负选择筛选,即构建一个含同靶基因序列的载体,对其进行标记,利用不同的Cas12亚蛋白对其进行切割,以观察哪些Cas12亚型可以靶向并切割大肠杆菌中编码必需基因的DNA,而导致细菌的生存能力的降低,最终筛选出了Cas12f、Un1Cas12f1及Cas12j等,各种CRISPR-Cas亚型的活性表现出了不同特征,涵盖了对ssRNA和ssDNA的靶标与旁切,以及对dsDNA的切割特性,这揭示了Cas12蛋白在其进化路径上经历的性质变迁。
Cas12f(Cas14)家族普遍“迷你”,Cas12f1蛋白大小紧凑,尺寸仅在400~700 aa左右,gRNA包括一个37 nt的crRNA和一个140 nt的tracrRNA,这可能与Cas12f1的ssDNA切割活性有关。最初发现Cas12f是一种靶向ssDNA的CRISPR内切酶,不需要PAM来激活[56,58]。如Un1Cas12f1(529 aa)和AsCas12f1(422 aa)已被证明在哺乳动物细胞中实现了强大的基因修饰和调控[51,57, 59-61]。Huang等[62]发现作为CRISPRa激活剂,Un1Cas12f1可诱导基因表达,其水平与Cas12a和Cas9相当,而作为DNA裂解编辑器,Cas12f1表现出与Cas12a类似的特性,如高特异性和主导诱导缺失而不是插入,但活性较低。Cas12j的独特之处在于:其用于使crRNA加工成熟和切割靶向DNA的单一活性位点的双重用途。这种多任务结构域在一定程度上解释了Cas12j整体尺寸的减小。Wang等[53]对Cas12j的六种亚型核酸酶进行了功能筛选,并确定Cas12j-8是一种理想的基因组编辑器,具有超紧凑的尺寸,该课题组通过实验证明,Cas12j-8能够对具有单核苷酸多态性(SNP)的基因进行等位基因特异性破坏。Cas12j-8能识别一个简单的5′-TTN-3′ PAM,提供了高目标位点密度。通过计算机程序模拟分析数据后显示,Cas12j-8不仅能对ClinVar数据库中的25 931个临床相关变体进行等位基因破坏,还能对dbSNP数据库中的485 130 147个SNP进行破坏。因此,Cas12j-8将特别适用于治疗性应用。它们的发现与鉴定为分子生物学、生物医学研究和临床治疗提供了新的工具。
2.2 蛋白工程化改造
IS200/IS605转座子家族中广泛存在的TnpB蛋白最近成为最小的RNA引导的核酸酶,能够在真核细胞中进行靶向基因组编辑[38, 63]。其中在2023年Xiang等 [64-65]的最新研究中表明TnpB是一种紧凑型RNA引导的DNA核酸内切酶,它是Cas12核酸酶的前体,其与Cas9一起被广泛用于基因组定向操作,能在人类数十种基因组位点上表现出强大的编辑能力,同时小的体积也为更多的基因编辑提供可能性[1, 66]。在这里,Sasnauskas等[67]揭示了TnpB核酸酶的基本结构以及得到生化实验支持的DNA目标识别和裂解的分子机制,结果表明TnpB代表了Cas12蛋白家族的最小结构和功能核心,并为开发基于TnpB的基因组编辑工具提供了一个框架。
Xu等[55]通过产生Cas12f变体(CasMINI)与转录激活剂融合,即引导RNA和蛋白质工程从V-F型Cas12f(Cas14)系统中创造了一个高效的微型Cas系统(CasMINI),其蛋白质大小仅为597 aa,同时仍然具有有效的DNA靶向能力。CasMINI可以驱动高水平的基因激活(最多可增加数千倍),而天然的Cas12f系统却无法在哺乳动物细胞中发挥作用。结果表明,CasMINI系统在基因激活方面具有与Cas12a相当的活性,具有高度的特异性,并能进行强大的碱基编辑和基因编辑。未来CasMINI能广泛用于体内外的细胞工程和基因治疗应用。dCasMINI-V4-VPR约为3.2 kb,dCas12a-VPR有5.3 kb,dCasMINI为AAV的递送带来了无限可能,此外研究还表明CasMINI系统可实现单碱基编辑与精准基因编辑。CasMINI切割基因组并经过NHEJ修复后可带来大片段(长达20 nt)的丢失,或许可带来大片段移除的应用前景。
2.3 gRNA改造
目前发现的最紧凑的CRISPR相关核酸酶之一是古细菌Un1Cas12f1。然而,Un1Cas12f1及其变体在真核细胞中的活性非常低。Kim等[60]根据Un1Cas12f1/gRNA/DNA的复合体结构信息,通过gRNA的系统改造,成功开发出迷你版CRISPR系统Cas12f1。截去了结构中定义为无序的RNA部分,同时在crRNA末端添加了富U序列;重新设计了Un1Cas12f1的天然引导RNA的五个部位:反式激活CRISPR RNA(tracrRNA)的5′端,tracrRNA-crRNA互补区,一个五(尿苷酸)序列,crRNA的3′端和tracrRNA中一个无序的茎2区。改造后的tracrRNA和crRNA极大增强了Un1Cas12f1在哺乳动物细胞中的基因编辑能力,在通过质粒载体、PCR扩增子和AAV传递时,能够在人类细胞中进行高效、特异的基因组编辑。由于Un1Cas12f1在原生质粒之外进行裂解,它可以被用来有效地创建大的缺失。工程化的Un1Cas12f1系统显示出与SpCas9相当的效率和与AsCas12a相似的特异性。目前,其余针对Cas12的gRNA改造在实验中尚未取得满足小型化需求的进展,因此关于该工具系统的整体分子量缩小的研究结果并不明显,在此方面,我们可以进行进一步探究。
整合一个小型化的Cas12和一个更短或优化过的gRNA可以有效避开载体大小的限制,对于基因治疗是有益的[68],可能会提高整个系统的交付效率,尤其对于难以转染的细胞或组织。对gRNA的改造和优化可以提高小型化Cas12的编辑效率、特异性和适用性,为基因编辑技术提供更强大和多样的工具。
3 Cas13的小型化研究
与目前广泛应用的Cas9或Cas12 DNA编辑工具不同,Cas13是一种RNA引导的可编辑RNA靶向单效应系统,能够在转录水平上进行基因操作[69]。CRISPR-Cas13最初被鉴定为由A、B、C、D四大类组成[66, 70],目前已有更多Cas13亚蛋白被筛选出来[71]。与Cas12或Cas9不同的是,它缺少一个DNase结构域,负责裂解pre-crRNA和目标RNA的两个RNase催化袋分别独立地位于Helical-1和HEPN结构域上[72-73](图4)。表4总结了Cas13的小型化相关研究。
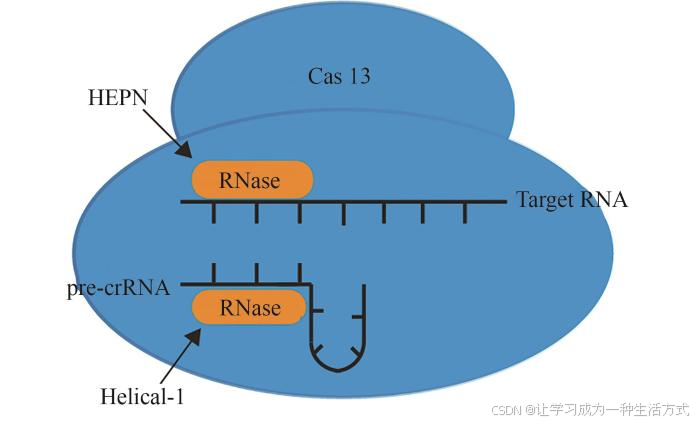
图4 Cas13结构示意图
Table 4 Miniaturization studies of Cas13
| 蛋白 | 分子量 /kDa | PFS | spacer /nt | tracrRNA | 直接重复 序列(DR) | ssRNA | 实验细胞/细菌 |
|---|---|---|---|---|---|---|---|
| Cas13a(C2c2)[74] | 138 | 3′-AT /C | 14~18 | N | 28 | Y | — |
| Cas13b(C2c6)[74-75] | 128 | 5′-D和3′-UAA | 30 | Y | 36 | Y | — |
| Cas13c(C2c7)[74] | 123 | 3′-U | 30 | N | 36 | Y | — |
| Cas13d[76] | 102 | N | 30 | N | 36 | Y | Ruminococcus flavefaciens FD-1 |
| Cas13Rx(RfxCas13d)[70] | 106 | 3′-A | 30 | N | 22 | Y | Ruminococcus flavefaciens FD-1 |
| Cas13X.1[77] | 85 | 5′-A/G/C-3′ | 30 | N | 36 | Y | HEK293T |
| Cas13Y.1[77] | 85 | N | 30 | N | 36 | Y | HEK293T |
| Cas13bt[78] | 85~88 | 5ʹ-D | 30 | N | 30 | Y | HEK293T |
| Cas13e3[71] | 84 | N | 27 | N | 30 | Y | — |
注:Y—C+T;D—A+G+T;N—A+C+G+T
3.1 筛选新型Cas13亚蛋白
Hu等[71]从测序数据中提取CRISPR阵列侧翼的20 kb DNA区域,用于预测开放阅读框(open reading frame,ORF)。选择具有超过400个残基的蛋白质进行进一步分析。随后构建了由NCBI数据库中所有已知Cas13a、Cas13b、Cas13c和Cas13d蛋白序列的隐马尔可夫模型(hidden Markov model, HMM)组成的Cas13库。使用这个包含所有已知Cas13蛋白特征的文库,可以最大限度地从未表征的蛋白质序列中发现潜在的新Cas13蛋白,并确定了5个新的Cas13分支:Cas13e至Cas13i,蛋白大小在740~1300 aa之间。新型Cas13系统的间隔区和DR的平均长度分别为30 nt和36 nt,其中,Cas13e3表现出与Cas13X.1和RfxCas13d相当的强大敲除活性。
Kannan等[78]在原核生物和病毒基因组以及元基因组中,从数千个Cas13酶中鉴定并表征了一个超小型Cas13酶——Cas13bt,其尺寸只有其他Cas13酶的一半大小,这种微型尺寸的RNA编辑工具可以被装进单个AAV中,从而实现体内的RNA编辑。研究通过使用腺苷和胞嘧啶脱氨酶Cas13bt进行功能化改造,设计了可用于AAV递送的紧凑型REPAIR和RESCUE结构,并实现了RNA编辑器在单个AAV中的包装,从而进一步推进可编程RNA编辑技术的发展。
3.2 蛋白工程化改造
Cas13蛋白的相对较大尺寸可能限制了其在某些应用中的效率,尤其是在病毒传递或细胞透过性方面。因此对于Cas13进行缩小化工程化改造以及优化尺寸和功能性能变得至关重要。Tang等[79]在研究中揭示了Cas13蛋白的主要结构域和其功能,通过定向进化,可以产生更小但活性不变甚至更高的Cas13变种,这些变种经过多轮筛选和优化,能在确保减小尺寸的同时保持所需的功能。Wang等[80]在研究中发现,通过截短或删除非关键的、不影响蛋白活性的区域,可以有效减小蛋白的尺寸。Li课题组[81]提出了一种基于蛋白质结构的蛋白质小型化IDC策略,在研究中发现了一种基于AlphaFold2预测的结构生成小型CRISPR-Cas13系统的可行性,这一方式使在保留蛋白质功能的同时最大限度地提高了蛋白质小型化,极大地促进了Cas13蛋白的体积缩减化发展。目前,通过各种策略,研究者们正在努力创建更小、更高效和更具功能性的新型Cas13,然而随着蛋白尺寸的减小,其稳定性和表达性可能会受到一定程度的影响。因此,对于小型化Cas13的结构进行进一步的优化,以确保其在生物系统中的有效表达和稳定性,是一个重要的研究方向。
3.3 gRNA改造
gRNA在Cas13中不仅能起到引导的作用,还能参与特异性识别。Bandaru等[82]的研究表明,Cas13的gRNA是一个多功能的组件,可以进行细致的调整,从而实现更广泛和高效的RNA目标操作。Xiao等[83]在实验中首先在培养细胞中筛选了几种以Myo6C442Y为靶标的基于Cas13的RNA碱基编辑器和gRNA变体,发现由截短的Cas13X.1和RNA编辑酶腺苷脱氨酶作用于RNA2脱氨酶结构域变体(ADAR2ddE488Q)组成的基于微型dCas13X.1的腺苷碱基编辑器(mxABE)表现出A>G转换效率高和脱靶编辑频率低的特点,并且经过小鼠体内验证发现,该CRISPR系统提高了含有目的基因细胞小鼠的存活率。这些研究表明,在优化gRNA的环境下,该系统的低脱靶率、高效率和明确的特异性可以进一步增强。这为利用gRNA的改进来减少Cas13总蛋白大小的实验策略提供了重要的理论基础。
4 结 语
综上所述,虽然小型化的CRISPR-Cas系统在理论和实验室研究中表现出了较大的优势,但在编辑活性、PAM限制、蛋白大小、特异性等方面仍存在一些问题[84]。譬如,传统的Cas9、Cas12a、Cas13a等核酸酶虽然具有良好的编辑功能,但它们因为蛋白分子量的大小而无法直接或同时携带其他的重要部件装进AAV里;若以其蛋白分子量大小为考量,选择性地缩减结构域或是编辑蛋白质结构后将其装入AAV中,又会在一定程度上大大限制基因编辑的效率和特异性。
随着科技的不断发展,CRISPR-Cas系统的小型化研究也日渐取得突破。通过筛选CRISPR亚蛋白后缩减结构域、改造gRNA、改造蛋白质结构等,以此获得的小型CRISPR-Cas系统,因其具有较小的蛋白质组成和较短的gRNA序列,而更易于最终的工程化和优化。随着基因组和宏基因组序列数据库的增加,将扩大对CRISPR生物学和核酸操作工具箱的理解。更高效、更迷你的CRISPR系统可以提高其基因编辑的效率和特异性,从而扩展CRISPR-Cas系统在基因编辑和治疗领域的应用[85]。这些改进亦促进了CRISPR-Cas技术在体内应用的all-in-one AAV传递载体的构建。当前,小型化的CRISPR-Cas系统在临床应用上尚未展现显著效果,其潜能仍待深入探索和开发。因此,我们需要进一步优化小型化的CRISPR-Cas系统,改进现有的基因编辑工具,使其在疾病治疗等领域发挥更大的作用。
对于CRISPR-Cas系统的小型化,另一个重要的研究方向是如何优化其在体内的传递。小型化的Cas蛋白可以更容易地穿透细胞膜和组织,可能具有更高的特异性和更少的副作用[86]。目前,已经有一些研究在尝试使用各种载体,如纳米粒子或脂质体,以提高CRISPR-Cas系统的体内稳定性和传递效率[87]。但是,这些方法可能会面临一些挑战,例如如何确保载体的稳定性和生物相容性,以及如何确保载体能有效地将CRISPR-Cas系统送到目标细胞。CRISPR-Cas系统的小型化研究是一个挑战和机遇并存的领域,我们需要更好地解析现有工具的作用机制、更有效地筛选新型蛋白体系等来达到CRISPR-Cas系统的合理小型化,这存在着许多挑战,但随着对CRISPR-Cas系统的理解不断深入,有理由相信,未来我们将能够开发出更小、更有效、更特异性的CRISPR-Cas系统,为基因编辑和基因治疗等领域带来更多的可能性。总之,小型化的CRISPR-Cas系统代表了一个引人注目的前沿领域,对基因编辑和治疗有着巨大的潜力。通过克服目前面临的技术挑战,我们可以期待看到CRISPR-Cas系统在临床应用中取得更大的成功,并为疾病治疗和基因编辑领域带来更多革命性的进展,如果能够继续支持和投资这一领域的研究,将能够有益于患者的福祉和健康。