易基因: Nat Plants:南科大朱健康/华中农大赵伦团队aChIP-seq+WGBS表观多组学揭示ROS1调控DNA去甲基化新机制
news2026/5/4 14:01:35
大家好这里是专注表观组学十余年领跑多组学科研服务的易基因。2026年4月2日华中农业大学赵伦教授与南方科技大学朱健康院士现澳门科技大学校长团队合作在《Nature Plants》期刊发表题为“Occupancy-based mechanism is the chief mode of ROS1 function in preventing DNA hypermethylation”的科研成果。该研究以甘蓝型油菜和拟南芥为对象系统揭示了DNA去甲基化酶ROS1在体内的主要作用机制打破此前ROS1主动去甲基化、从而抑制全基因组DNA高甲基化的传统认知提出并验证“占位机制Occupancy-based mechanism”是ROS1维持基因组DNA低甲基化的主要模式。本研究通过优化版染色质免疫共沉淀测序advanced ChIP-seqaChIP-seq和DNA甲基化组分析WGBS等表观多组学技术揭示ROS1通过其N端和C端结构域广泛结合几乎所有染色质开放区域通过物理占位拮抗RNA介导DNA甲基化RdDM通路的招募从而以“被动去甲基化”方式维持DNA低甲基化状态而非依赖广泛的酶切活性。此外研究还发现ROS1是染色质可及性的关键标志物和调控因子在DNA甲基化依赖和非依赖环境中均发挥染色质可及性调控作用并作为“储备保护者”或“现役保护者”动态响应表观遗传环境变化为作物表观遗传精准改良提供关键理论支撑。英文标题Occupancy-based mechanism is the chief mode of ROS1 function in preventing DNA hypermethylation译文标题基于占位机制是ROS1抑制DNA高甲基化的主要功能模式发表时间2026年4月2日发表期刊Nature Plants影响因子2025IF 13.6/Q1技术平台ChIP-seq、WGBS、ATAC-seq、RNA-seq等作者单位华中农业大学、南方科技大学等DOI10.1038/s41477-026-02258-z易小结本研究不仅重新定义ROS1在DNA甲基化调控和染色质可及性中的多重作用更揭示其通过“占位机制”实现基因组稳态的表观遗传调控机制为理解真核生物平衡表观遗传重编程与基因组完整性提供了新思路。研究应用aChIP-seq绘制了ROS1的全基因组结合图谱WGBS绘制了单碱基分辨率的DNA甲基化图谱是精准鉴定差异甲基化区域DMRs、定量甲基化水平变化、并最终验证“占位导致被动去甲基化”机制的重要方法。表观多组学技术联用在揭示复杂表观调控网络、解析作物农艺性状表观遗传基础等前沿研究中发挥重要作用。研究方法1、材料构建多种拟南芥突变体和转基因株系构建表达Flag标签的全长ROS1、截短变体如ΔN294、ΔC146以及催化失活突变体的转基因植物。甘蓝型油菜ROS1同源基因也采用类似策略。2、机制探索aChIP-seq优化版染色质免疫沉淀测序团队自主研发aChIP方法用于低丰度蛋白和特殊植物组织成功绘制ROS1及其家族蛋白DME/DML2/DML3以及RNA聚合酶IVNRPD1的全基因组高分辨率结合图谱。ATAC-seq染色质可及性测序全基因组范围内检测染色质可及性并分析其在不同基因型和组织中变化。全基因组重亚硫酸盐测序WGBS对野生型Col-0、ros1-4突变体、nrpd1-3、nrpd1-3 ros1-4以及各种转基因回补株系等进行WGBS分析。绘制全基因组DNA甲基化图谱、鉴定ros1-4中的差异高甲基化区域hyper-DMRs、比较不同基因型甲基化水平区分“主动去甲基化主导”和“被动去甲基化主导”位点、验证ROS1占位对抑制DNA甲基化的作用。RNA-seq转录组测序分析基因表达变化。组蛋白修饰ChIP-seq分析H3K4me3、H3K9ac、H3K4me1、H3K27me3和H3K9me2等代表性组蛋白修饰。3、体外酶活性实验验证ROS1及其D971A突变体的5mC糖基化酶/裂解酶活性。结果图形1ROS1占位几乎所有染色质开放区域研究首先通过优化版aChIP-seq技术成功绘制了ROS1的全基因组结合图谱。分析发现ROS1结合位点与ATAC-seq所定义的染色质开放区域高度共定位图1a-c。在不同组织幼苗地上部、根、茎和不同基因型组蛋白去乙酰化酶突变体hda6中ROS1占位水平与染色质可及性信号强正相关图1d-h。上述研究结果表明ROS1并非像先前经典研究认为的仅被招募到少数特定序列位点而是广泛地、非序列特异性地与几乎所有的染色质开放区域结合使其成为一个染色质可及性的关键标志物。这一发现是理解ROS1新功能机制的基础。图1ROS1占位机制随组织和基因型变化并与染色质可及性改变相关2N端和C端结构域对ROS1结合染色质可及性至关重要为探究ROS1如何结合染色质研究人员构建了敲除N端富集赖氨酸区域ΔN294、C端RNA识别基序结构域ΔC146或两者同时缺失ΔN294ΔC146的截短突变体。aChIP-seq分析显示这些截短显著减少甚至完全抑制ROS1与染色质可及性的结合力图2a-d。然而有趣的是这些仍能微弱结合的截短变体在全基因组范围内的分布模式与全长ROS1相似仍靶向避开异染色质标记H3K9me2区域。上述研究结果表明ROS1的N端和C端是其与染色质结合的主要决定因子而特定染色质环境如特定的组蛋白修饰可能对其结合有辅助作用。同时研究还发现ROS1家族其他成员DME/DML2/DML3以及甘蓝型油菜中的ROS1同源蛋白也表现出与染色质可及性相关的类似结合模式。图2N端和C端结构域对ROS1结合染色质可及性至关重要3ROS1直接调控仅部分ros1-4差异高甲基化区域鉴于ROS1在DNA去甲基化中的已知作用研究人员进一步探讨了ROS1占位与野生型植物中DNA甲基化的关系。WGBS分析显示DNA甲基化通常发生在ROS1结合位点之外而ROS1未结合位点则广泛发生甲基化图3a表明ROS1占位与DNA甲基化呈负相关。为检验ROS1在其结合位点维持低甲基化的作用研究人员通过整合ROS1结合谱和WGBS鉴定的ros1-4突变体中的差异高甲基化区域hyper-DMRs将基因组区域分为三类图3bI型区域ROS1结合但未发生高甲基化占ROS1结合位点的94%、II型区域ROS1结合且发生高甲基化占hyper-DMRs的25%和III型区域未检测到ROS1结合但发生高甲基化占hyper-DMRs的75%。这一分类清晰表明在绝大多数94%ROS1结合位点即使ROS1缺失也维持DNA低甲基化说明ROS1对于维持这些位点低甲基化并非必需。ROS1直接抑制DNA高甲基化的作用仅限于其结合位点中一小部分25%在ros1-4中的高甲基化区域II型。而大部分75%高甲基化区域III型未与ROS1结合ROS1通过间接机制抑制DNA高甲基化。图3ROS1占位、RdDM招募与DNA甲基化之间的关系4ROS1占位通过拮抗RdDM招募以维持DNA低甲基化为阐明ROS1如何直接维持低甲基化研究聚焦于RNA介导的DNA甲基化RdDM通路。通过分析ROS1和RdDM核心组件Pol IVNRPD1结合谱并综合分析ros1-4、nrpd1-3、nrpd1-3 ros1-4和野生型的DNA甲基化数据研究人员发现两种不同类型的RdDM靶位点图3fI型位点经典靶点无ROS1结合始终高甲基化和II型位点拮抗位点在野生型中ROS1占位而Pol IV信号和DNA低甲基化在ros1-4中ROS1缺失导致Pol IV招募和DNA甲基化显著增加图3e-i。为鉴定ROS1占位发挥功能的位点研究者通过连续基因靶向方法生成ROS1原位D971A替换植株并分析其相对于野生型和ros1-4的甲基化谱。分析结果发现在鉴定的1,444个ROS1结合hyper-DMRsros1-4 vs Col-0中93%保持与野生型相似的低甲基化状态仅7%102个位点表现出与ros1-4一致的强高甲基化图3j-k。这些结果进一步支持ROS1主要通过染色质占位机制在大多数靶位点维持低甲基化。5ROS1占位介导广泛非酶活性依赖的被动DNA去甲基化体外酶活性实验已显示ROS1作为糖基化酶/裂解酶活性glycosylase/lyase酶切5mC碱基以启动主动DNA去甲基化。但ROS1在体内通过主动或被动机制介导DNA去甲基化仍然未知。ROS1占位通过拮抗甲基化以维持低甲基化状态图3i这引出了其通过酶活性非依赖性方式介导被动去甲基化的假设。研究人员构建了催化活性位点突变体ROS1D971A酶活性缺失但结合能力保留。将野生型ROS1和ROS1D971A分别回补到ros1-4突变体中并进行DNA甲基化比较图4。研究鉴定出一批在ros1-4中高甲基化、能被野生型ROS1回补降低甲基化的位点。约65%的这些位点也能被缺失酶活性的ROS1D971A回补降低甲基化被动去甲基化主导位点而另一部分则不能主动去甲基化主导位点。更重要的是在ROS1D971A原位替换的植物中约94%的ROS1靶位点维持DNA低甲基化。这些发现表明ROS1占位介导的被动DNA去甲基化在植物中广泛发生。图4ROS1占位介导不依赖糖基化酶/裂解酶活性的被动DNA去甲基化。体外糖基化酶/裂解酶实验显示ROS1中Asp971突变为丙氨酸可抑制其5mC切割活性。野生型全长ROS1与突变体ROS1D971A-Flag/ros1-4在所有24,226个可及染色质区域峰周围区域的占位关联。ROS1 D971A表现出与野生型ROS1相似的全基因组占位和表观基因组特征。以野生型ROS1去甲基化位点为对照鉴定ros1-4中ROS1D971A去甲基化位点。共鉴定出603个野生型ROS1去甲基化位点这些位点被野生型ROS1和突变体ROS1D971A共同占位左图。其中396个位点在至少一个独立的ROS1D971A-Flag/ros1-4株系中被ROS1D971A去甲基化归为被动主导位点右图。剩余位点归为主动主导位点。主动主导ROS1D971A未去甲基化和被动主导ROS1D971A去甲基化位点的DNA甲基化水平。d和e的代表性示例。ChIP-seq信号以CPM显示甲基化水平以甲基化胞嘧啶比例显示。6ROS1独立或协同其他表观遗传调控因子维持染色质开放既然ROS1占位开放染色质研究进一步探究它是否影响可及性本身。ATAC-seq分析发现在ros1-4中有597个区域的可及性降低LARs而可及性升高区域HARs较少仅10个图5a-b表明ROS1能独立调控一部分染色质开放性。在rdd-2和drdd等多突变体中发现更多可及性降低区域且部分与这些突变体特有的hyper-DMRs重叠表明ROS1与同源蛋白DME、DML2/3在功能上存在冗余。此外与组蛋白乙酰转移酶GCN5或甲基转移酶SDG2的双突变分析也揭示ROS1与这些活性染色质修饰因子之间存在协同或冗余的多重调控机制共同维护关键区域的染色质开放性图5c-d。图5ROS1独立或与GCN5冗余维持染色质可及性7ROS1作为特定基因组区域染色质可及性的“储备保护者”发挥作用鉴于ROS1移除后大量开放区域持续存在研究者提出ROS1可能作为其中部分区域的储备保护者。为验证此假设尝试通过突变DNA甲基化调控因子建立有无DNA甲基化的比较系统然后评估ROS1移除后的染色质可及性变化图6a。在拟南芥中DNA甲基化主要由RdDM通路介导由五个功能性DNA甲基转移酶维持其中MET1负责CG甲基化DRM1、DRM2、CMT2和CMT3协同负责非CG甲基化。然而这些通路突变几乎完全抑制ROS1启动子DNA甲基化导致ROS1表达显著下调。而HDA6则通过与MET1互作调控特定位点DNA甲基化。尽管hda6突变体DNA甲基化降低且ROS1表达下调但仍有ROS1表达。因此研究人员使用hda6突变体建立有HDA6依赖DNA甲基化ros1-4 vs野生型和无HDA6依赖DNA甲基化hda6 ros1-4 vs hda6的比较系统评估ROS1移除后的染色质可及性变化图6b。结果共鉴定出290个无论HDA6存在与否ROS1均稳定结合的位点图6c-d。在存在HDA6依赖DNA甲基化时ros1-4 vs野生型ROS1移除导致DNA甲基化沉积和染色质可及性降低证明ROS1在这些位点以DNA甲基化依赖方式维持染色质开放而在无HDA6依赖DNA甲基化时hda6 ros1-4 vs hda6即使移除ROS1这些位点仍保持开放图6b,d。因此ROS1广泛结合染色质可及性但并非在所有结合位点发挥功能而是作为“储备保护者”仅在DNA甲基化发生时才变为“现役保护者”。图6ROS1在特定基因组区域负责染色质可及性的储备保护者8ROS1在DNA甲基化依赖和非依赖环境中调控染色质可及性鉴于ROS1在调控染色质可及性和DNA甲基化中的重要作用研究人员接下来分析了ROS1在特定位点调控染色质可及性的作用是否依赖DNA甲基化。研究对ros1-4中597个可及性降低区域LARs的DNA甲基化数据分析显示图7a其中543个位点在ros1-4中DNA甲基化水平显著上升DNA甲基化依赖性调控而54个位点在两种基因型中甲基化水平都很低DNA甲基化非依赖性调控。进一步分析发现一部分DNA甲基化依赖性的LARs与前述的RdDM II型位点重叠58个图7b。这些位点ROS1移除导致的可及性下降在nrpd1-3 ros1-4双突变中因RdDM通路被同时破坏而得以恢复图7c证实了“ROS1占位→拮抗RdDM→维持低甲基化和开放性”轴的存在。同时组织间的可及性动态变化大多发生在DNA甲基化缺失区域图7e表明并非所有染色质可及性变化都由DNA甲基化介导。图7ROS1在DNA甲基化依赖和非依赖环境中调控染色质可及性结论和启示本研究通过aChIP-seq、WGBS、ATAC-seq、RNA-seq等表观多组学综合分析揭示了ROS1主要基于其蛋白对染色质开放区域的物理占位以抑制DNA高甲基化的表观遗传调控机制。ROS1通过拮抗DNA甲基化复合体招募以促进被动去甲基化而非依赖其糖基化酶/裂解酶活性主动碱基切除的经典认知。机制创新ROS1主要通过“占位机制”而非酶催化活性维持DNA低甲基化破解了全基因组碱基切除与基因组稳定性之间传统认知的全新机制。功能拓展ROS1是染色质可及性的关键标志物和调控因子在DNA甲基化依赖和非依赖环境中均发挥调控作用。动态调控ROS1可作为“储备保护者”或“主动保护者”动态响应表观遗传环境变化。技术参考aChIP和WGBS等技术的整合应用为低丰度表观调控因子的全基因组研究提供了方法学参考。参考文献Deng L, Zhu G, Zhong W, Jia Z, Zhou Q, Zhang M, Si Z, Zhang Q, Liang Y, Du X, Mao Y, Wu J, Li G, Shen J, Zhu JK, Zhao L. Occupancy-based mechanism is the chief mode of ROS1 function in preventing DNA hypermethylation. Nat Plants. 2026 Apr 2. doi: 10.1038/s41477-026-02258-z.
本文来自互联网用户投稿,该文观点仅代表作者本人,不代表本站立场。本站仅提供信息存储空间服务,不拥有所有权,不承担相关法律责任。如若转载,请注明出处:http://www.coloradmin.cn/o/2538926.html
如若内容造成侵权/违法违规/事实不符,请联系多彩编程网进行投诉反馈,一经查实,立即删除!相关文章
SpringBoot-17-MyBatis动态SQL标签之常用标签
文章目录 1 代码1.1 实体User.java1.2 接口UserMapper.java1.3 映射UserMapper.xml1.3.1 标签if1.3.2 标签if和where1.3.3 标签choose和when和otherwise1.4 UserController.java2 常用动态SQL标签2.1 标签set2.1.1 UserMapper.java2.1.2 UserMapper.xml2.1.3 UserController.ja…
wordpress后台更新后 前端没变化的解决方法
使用siteground主机的wordpress网站,会出现更新了网站内容和修改了php模板文件、js文件、css文件、图片文件后,网站没有变化的情况。
不熟悉siteground主机的新手,遇到这个问题,就很抓狂,明明是哪都没操作错误&#x…
网络编程(Modbus进阶)
思维导图 Modbus RTU(先学一点理论)
概念 Modbus RTU 是工业自动化领域 最广泛应用的串行通信协议,由 Modicon 公司(现施耐德电气)于 1979 年推出。它以 高效率、强健性、易实现的特点成为工业控制系统的通信标准。 包…
UE5 学习系列(二)用户操作界面及介绍
这篇博客是 UE5 学习系列博客的第二篇,在第一篇的基础上展开这篇内容。博客参考的 B 站视频资料和第一篇的链接如下:
【Note】:如果你已经完成安装等操作,可以只执行第一篇博客中 2. 新建一个空白游戏项目 章节操作,重…
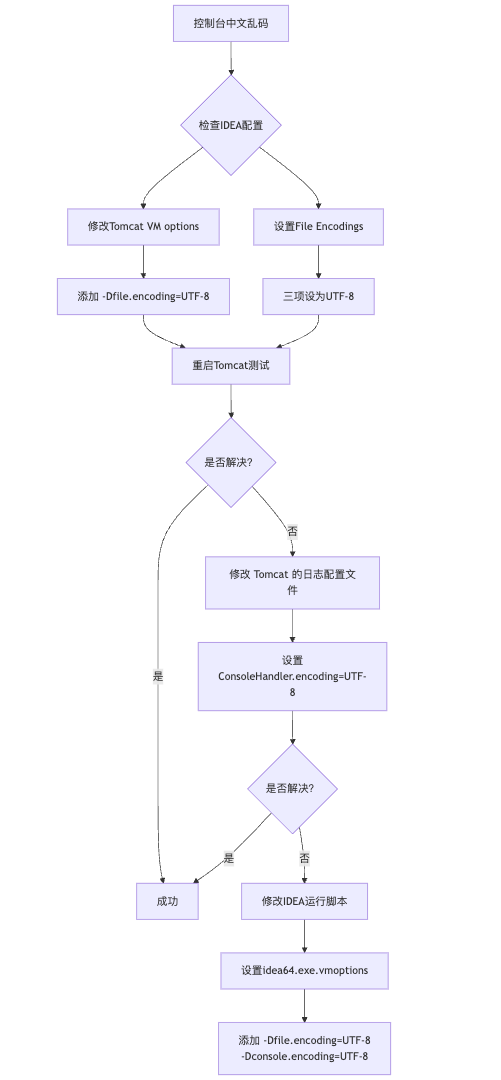
IDEA运行Tomcat出现乱码问题解决汇总
最近正值期末周,有很多同学在写期末Java web作业时,运行tomcat出现乱码问题,经过多次解决与研究,我做了如下整理:
原因:
IDEA本身编码与tomcat的编码与Windows编码不同导致,Windows 系统控制台…

利用最小二乘法找圆心和半径
#include <iostream>
#include <vector>
#include <cmath>
#include <Eigen/Dense> // 需安装Eigen库用于矩阵运算 // 定义点结构
struct Point { double x, y; Point(double x_, double y_) : x(x_), y(y_) {}
}; // 最小二乘法求圆心和半径 …
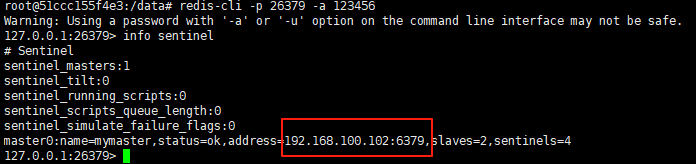
使用docker在3台服务器上搭建基于redis 6.x的一主两从三台均是哨兵模式
一、环境及版本说明
如果服务器已经安装了docker,则忽略此步骤,如果没有安装,则可以按照一下方式安装: 1. 在线安装(有互联网环境): 请看我这篇文章 传送阵>> 点我查看 2. 离线安装(内网环境):请看我这篇文章 传送阵>> 点我查看
说明:假设每台服务器已…
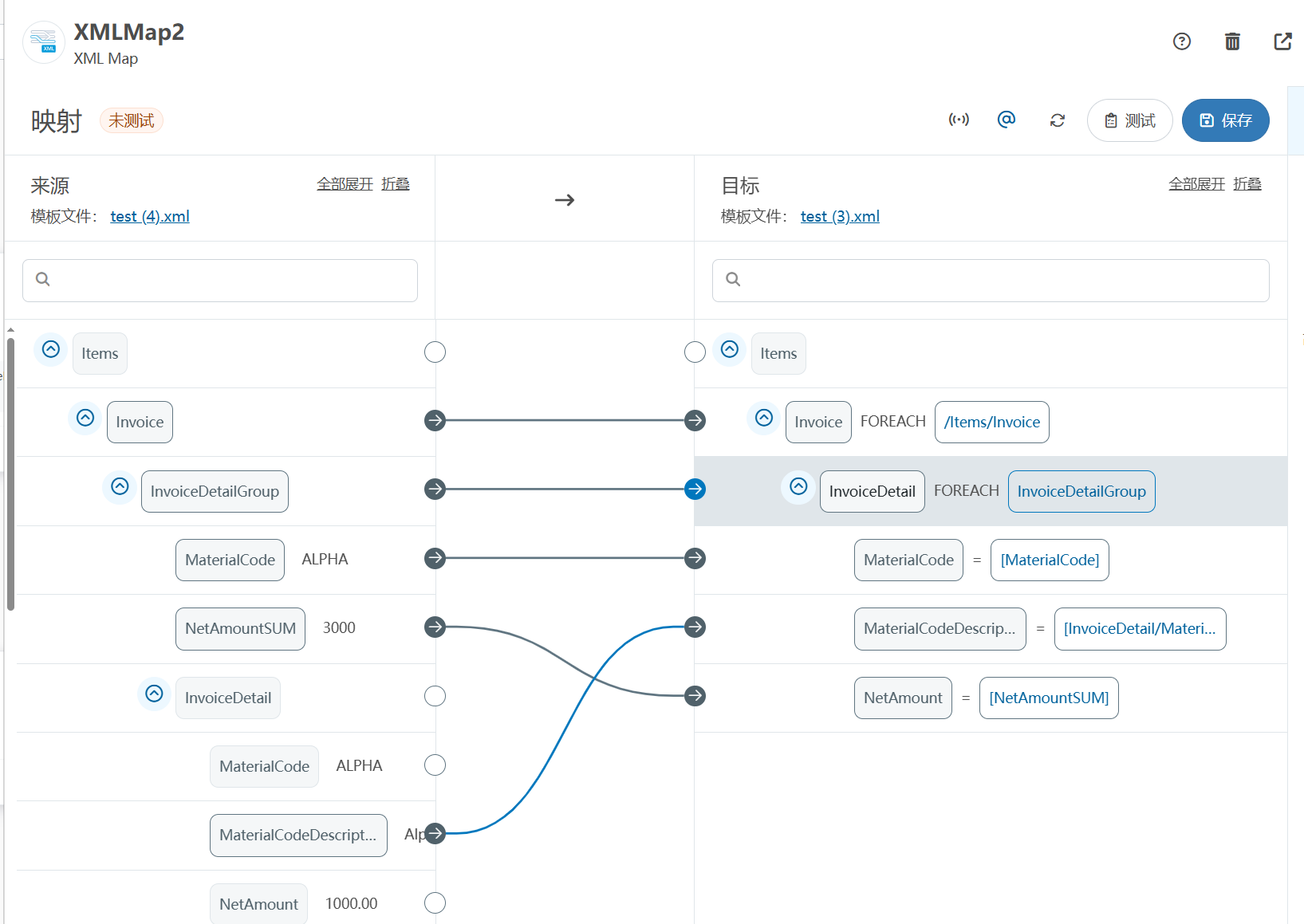
XML Group端口详解
在XML数据映射过程中,经常需要对数据进行分组聚合操作。例如,当处理包含多个物料明细的XML文件时,可能需要将相同物料号的明细归为一组,或对相同物料号的数量进行求和计算。传统实现方式通常需要编写脚本代码,增加了开…
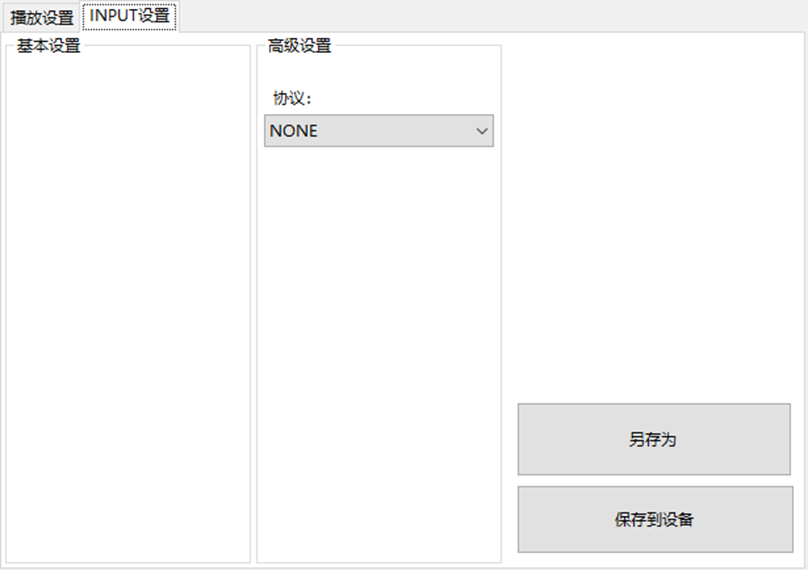
LBE-LEX系列工业语音播放器|预警播报器|喇叭蜂鸣器的上位机配置操作说明
LBE-LEX系列工业语音播放器|预警播报器|喇叭蜂鸣器专为工业环境精心打造,完美适配AGV和无人叉车。同时,集成以太网与语音合成技术,为各类高级系统(如MES、调度系统、库位管理、立库等)提供高效便捷的语音交互体验。
L…

(LeetCode 每日一题) 3442. 奇偶频次间的最大差值 I (哈希、字符串)
题目:3442. 奇偶频次间的最大差值 I 思路 :哈希,时间复杂度0(n)。 用哈希表来记录每个字符串中字符的分布情况,哈希表这里用数组即可实现。
C版本:
class Solution {
public:int maxDifference(string s) {int a[26]…

【大模型RAG】拍照搜题技术架构速览:三层管道、两级检索、兜底大模型
摘要
拍照搜题系统采用“三层管道(多模态 OCR → 语义检索 → 答案渲染)、两级检索(倒排 BM25 向量 HNSW)并以大语言模型兜底”的整体框架: 多模态 OCR 层 将题目图片经过超分、去噪、倾斜校正后,分别用…
【Axure高保真原型】引导弹窗
今天和大家中分享引导弹窗的原型模板,载入页面后,会显示引导弹窗,适用于引导用户使用页面,点击完成后,会显示下一个引导弹窗,直至最后一个引导弹窗完成后进入首页。具体效果可以点击下方视频观看或打开下方…
接口测试中缓存处理策略
在接口测试中,缓存处理策略是一个关键环节,直接影响测试结果的准确性和可靠性。合理的缓存处理策略能够确保测试环境的一致性,避免因缓存数据导致的测试偏差。以下是接口测试中常见的缓存处理策略及其详细说明:
一、缓存处理的核…
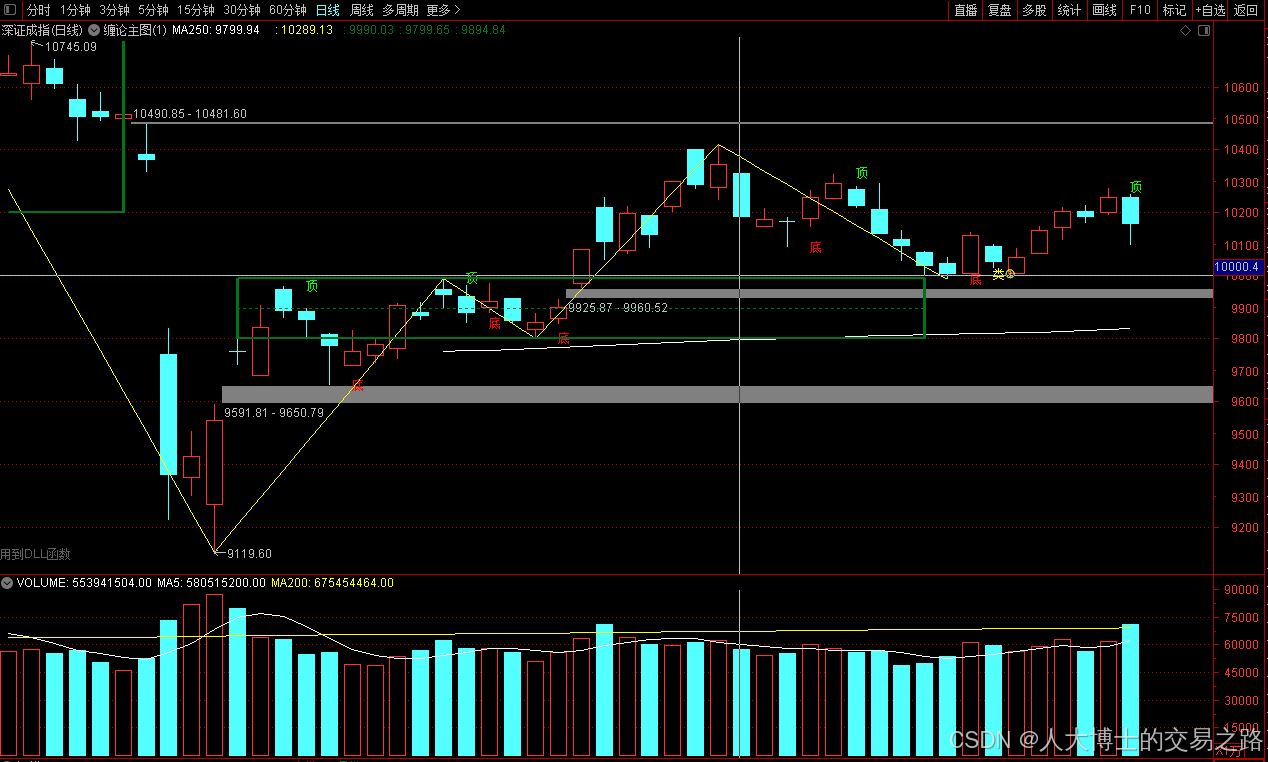
龙虎榜——20250610
上证指数放量收阴线,个股多数下跌,盘中受消息影响大幅波动。 深证指数放量收阴线形成顶分型,指数短线有调整的需求,大概需要一两天。 2025年6月10日龙虎榜行业方向分析 1. 金融科技
代表标的:御银股份、雄帝科技
驱动…
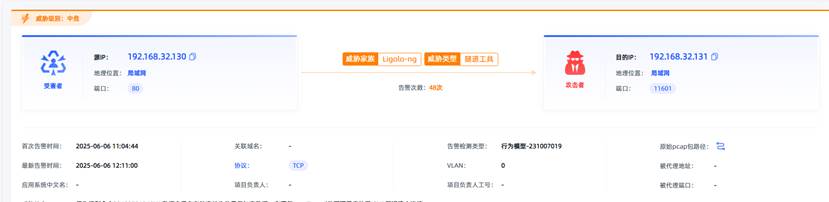
观成科技:隐蔽隧道工具Ligolo-ng加密流量分析
1.工具介绍
Ligolo-ng是一款由go编写的高效隧道工具,该工具基于TUN接口实现其功能,利用反向TCP/TLS连接建立一条隐蔽的通信信道,支持使用Let’s Encrypt自动生成证书。Ligolo-ng的通信隐蔽性体现在其支持多种连接方式,适应复杂网…

铭豹扩展坞 USB转网口 突然无法识别解决方法
当 USB 转网口扩展坞在一台笔记本上无法识别,但在其他电脑上正常工作时,问题通常出在笔记本自身或其与扩展坞的兼容性上。以下是系统化的定位思路和排查步骤,帮助你快速找到故障原因:
背景:
一个M-pard(铭豹)扩展坞的网卡突然无法识别了,扩展出来的三个USB接口正常。…

未来机器人的大脑:如何用神经网络模拟器实现更智能的决策?
编辑:陈萍萍的公主一点人工一点智能 未来机器人的大脑:如何用神经网络模拟器实现更智能的决策?RWM通过双自回归机制有效解决了复合误差、部分可观测性和随机动力学等关键挑战,在不依赖领域特定归纳偏见的条件下实现了卓越的预测准…

Linux应用开发之网络套接字编程(实例篇)
服务端与客户端单连接
服务端代码
#include <sys/socket.h>
#include <sys/types.h>
#include <netinet/in.h>
#include <stdio.h>
#include <stdlib.h>
#include <string.h>
#include <arpa/inet.h>
#include <pthread.h>
…

华为云AI开发平台ModelArts
华为云ModelArts:重塑AI开发流程的“智能引擎”与“创新加速器”!
在人工智能浪潮席卷全球的2025年,企业拥抱AI的意愿空前高涨,但技术门槛高、流程复杂、资源投入巨大的现实,却让许多创新构想止步于实验室。数据科学家…

深度学习在微纳光子学中的应用
深度学习在微纳光子学中的主要应用方向
深度学习与微纳光子学的结合主要集中在以下几个方向:
逆向设计 通过神经网络快速预测微纳结构的光学响应,替代传统耗时的数值模拟方法。例如设计超表面、光子晶体等结构。
特征提取与优化 从复杂的光学数据中自…