iMeta | 浙江农科院卢立志/曾涛联合中南大学湘雅医院揭示人参皂苷Rg3缓解肝脏铁死亡的新机制
news2026/4/2 11:00:22
点击蓝字 关注我们一种生物活性人参皂苷改善非酒精性脂肪性肝炎中氧化磷脂积累引起的肝细胞铁死亡iMeta主页http://www.imeta.science研究论文● 原文:iMeta(IF 33.2, 中科院双一区Top)●英文题目: A bioactive ginsenoside alleviates hepatocellular ferroptosis induced by oxidized phospholipid accumulation in nonalcoholic steatohepatitis●中文题目: 一种生物活性人参皂苷改善非酒精性脂肪性肝炎中氧化磷脂积累引起的肝细胞铁死亡● 原文链接: https://onlinelibrary.wiley.com/doi/10.1002/imt2.70114● DOI: https://doi.org/10.1002/imt2.70114● 2026年3月3日浙江省农业科学院卢立志、曾涛与中南大学湘雅医院史达尊等在iMeta发表了题 “A bioactive ginsenoside alleviates hepatocellular ferroptosis induced by oxidized phospholipid accumulation in nonalcoholic steatohepatitis” 的文章。● 本研究通过细胞实验、动物模型及临床样本验证等系统阐明了20(S)-人参皂苷Rg320 (S)-Rg3通过调控SPOP/ATF3/NUPR1信号轴抑制氧化磷脂OxPLs积累和肝细胞铁死亡从而缓解非酒精性脂肪性肝炎NASH的分子机制为NASH的靶向治疗提供了全新策略和候选药物分子。● 第一作者宗一博● 通讯作者曾涛zengtao4009126.com、卢立志lulizhibox163.com、史达尊shidazuncsu.edu.cn● 合作作者龙果、顾天天、黄攀、田勇、许文武、胡希恒● 主要单位浙江省农业科学院、中南大学湘雅医院、江苏大学亮 点●氧化磷脂OxPLs积累会推动非酒精性脂肪性肝炎NASH的发生发展而 20 (S)- 人参皂苷 Rg320 (S)-Rg3可显著降低小鼠体内的 OxPLs 含量● E3 泛素连接酶 SPOP 可介导转录因子 ATF3 的 K48 位泛素化修饰进而促进 ATF3 的蛋白酶体依赖性降解● 20 (S)-Rg3 通过调控 SPOP/ATF3/NUPR1 信号轴发挥抗肝细胞铁死亡作用Rg3 激活 SPOP 后加速 ATF3 降解解除其对线粒体稳态核心调控因子 NUPR1 的转录抑制最终恢复线粒体功能、减少脂质过氧化改善铁死亡。摘 要氧化磷脂OxPLs积累与铁死亡在非酒精性脂肪性肝炎NASH疾病中扮演关键角色但两者在NASH中的因果关联及调控机制仍不清楚。我们的前期研究结果表明20 (S) -人参皂苷 Rg320 (S)-Rg3可有效改善笼养应激诱导的蛋鸭肝脏氧化损伤以及炎症为了探究调控机制我们进一步开展了小鼠实验。本研究中活性人参皂苷20(S)-Rg3可显著缓解高脂饮食诱导的小鼠NASH症状包括改善血脂紊乱、肝脏炎症、纤维化及胰岛素抵抗。机制上Rg3可结合并激活E3泛素连接酶SPOP促进转录因子ATF3的K48位泛素化降解染色质免疫沉淀测序ChIP-seq以及双荧光素酶报告基因实验证实ATF3可结合Nupr1启动子区域并抑制其转录而Nupr1作为线粒体稳态的核心调控因子其表达下调会加剧线粒体氧化应激和铁死亡。Rg3通过降解ATF3解除对Nupr1的转录抑制恢复线粒体功能减少OxPLs积累和脂质过氧化最终抑制肝细胞铁死亡。临床样本分析显示肝硬化患者肝组织中SPOP和Nupr1表达下调ATF3和OxPLs水平升高提示该信号轴与人类肝脏疾病密切相关。综上本研究揭示了Spop-Atf3-Nupr1轴在NASH铁死亡中的关键调控作用证实Rg3是靶向该通路的潜在治疗药物同时为NASH的诊断提供了OxPLs这一无创生物标志物。视频解读Bilibilihttps://www.bilibili.com/video/BV1sxXUB5EVe/Youtubehttps://youtu.be/8E6DZibxhqc中文翻译、PPT、中/英文视频解读等扩展资料下载请访问期刊官网http://www.imeta.science/全文解读引 言非酒精性脂肪性肝炎NASH已成为慢性肝病的主要病因但目前尚无获批的治疗药物阐明其发病机制并寻找有效治疗药物迫在眉睫。人参皂苷Rg3已被报道具有肝脏保护作用但Rg3改善NASH的具体功能及分子机制尚未明确。氧化磷脂OxPLs作为生物活性脂质介质在代谢疾病中具有促炎和促动脉粥样硬化的特性可通过调控细胞信号通路和转录程序诱导细胞应激和死亡。越来越多的证据表明氧化应激可通过OxPLs积累促进铁死亡。已有研究表明在多种NASH小鼠模型和患者的肝脏组织及循环中OxPLs水平显著升高但由于OxPLs主要来源于非酶促脂质过氧化目前缺乏有效的中和策略其在NASH疾病进程中的具体作用与致病机制仍不明确。铁死亡是一种铁依赖的调控性细胞死亡可由磷脂过氧化驱动其特征为铁积累和脂质过氧化物形成。已有研究表明铁死亡参与NASH的病理过程但OxPLs积累与铁死亡之间的调控网络有待进一步的探究。本研究推测20(S)-人参皂苷Rg3可能通过调控OxPLs代谢和铁死亡相关通路发挥NASH保护作用。为验证这一假设我们通过高脂饮食诱导的NASH小鼠模型、OxPAPC诱导的肝细胞铁死亡模型结合临床样本分析系统探究了Rg3的作用及分子机制旨在为NASH的治疗提供新的理论依据和候选药物。结 果高剂量Rg3改善高脂饮食诱导的小鼠NASH为评估Rg3的治疗潜力我们通过给小鼠喂食高脂饮食24周建立NASH模型图1A。高剂量Rg3显著降低小鼠体重、肝脏重量及肝体比同时改善血脂紊乱表现为小鼠肝脏甘油三酯TG、总胆固醇TC、非酯化脂肪酸NEFA和低密度脂蛋白胆固醇LDL-C水平降低图1B、C、S1A-E。高剂量Rg3还降低了血浆丙氨酸转氨酶ALT、天冬氨酸转氨酶AST和乳酸脱氢酶LDH水平图1D提示其对肝脏损伤的保护作用。组织病理学分析证实Rg3显著减少肝细胞气球样变、脂肪变性和纤维化图1E、S1F-I。高剂量Rg3通过抑制肝脏组织和血浆中的关键促炎细胞因子TNFα、IL-6、IL-1β发挥抗炎作用图S1J-O。鉴于NASH与全身代谢紊乱密切相关我们进一步评估了整体葡萄糖稳态发现高剂量Rg3显著改善小鼠的葡萄糖耐量和胰岛素敏感性图1F-I。基于以上研究结果后续机制探究均采用高剂量Rg3以下简称Rg3。Rg3缓解铁死亡并上调肝脏SPOP表达转录组和蛋白组学分析结果一致表明铁死亡是高脂饮食诱导NASH中富集的重要致病通路之一而Rg3处理可显著抑制该通路图1J、S2A-C。Rg3改善了高脂饮食导致的肝脏氧化损伤表现为过氧化氢酶CAT和超氧化物歧化酶SOD活性增加谷胱甘肽GSH水平和GSH/氧化型谷胱甘肽GSSG比值升高丙二醛MDA和GSSG水平降低图S2D-I。Rg3显著减轻高脂饮食小鼠的肝脏铁沉积图1K、L、S2J。值得注意的是与健康对照组相比纤维化小鼠和肝硬化患者肝脏中的总OxPAPC氧化磷脂的混合物水平均显著升高图1M、S2K。靶向HPLC-MS脂质组学进一步显示Rg3处理后多种OxPLs含量降低图1N、S2L。同时Rg3显著降低肝脏中脂质过氧化和铁死亡的经典标志物4-羟基壬烯醛4-HNE水平并减轻高脂饮食诱导的线粒体损伤图S2M-N。整合多组学数据后我们将E3泛素连接酶SPOP鉴定为关键候选分子图1O。SPOP已被证实通过抑制Tlr4/Nf-κb通路发挥抗炎作用并在肝细胞癌中作为肿瘤抑制因子但在铁死亡中的作用尚不明确。结果显示Rg3显著上调了高脂饮食小鼠肝脏中Spop在mRNA和蛋白水平的表达图1P、S2O、P提示SPOP可能是Rg3抑制铁死亡的关键调控因子。图1. 高剂量人参皂苷Rg3可改善高脂饮食HFD诱导的小鼠非酒精性脂肪性肝炎NASH并上调肝脏中Spop基因的表达(A) Rg3干预策略示意图用于HFD诱导的NASH小鼠模型。(B) 对照组与Rg3处理组小鼠的体重、肝脏重量及肝重/体重比值LW/BWn 8。(C) 肝脏中甘油三酯TG、总胆固醇TC及非酯化脂肪酸NEFA水平n 8。(D) 血浆中丙氨酸氨基转移酶ALT、天冬氨酸氨基转移酶AST及乳酸脱氢酶LDH活性n 8。(E) 肝组织切片代表性苏木精-伊红HE、油红OORO、天狼星红PSR及IV型胶原CIV染色结果。(F, G) 小鼠腹腔注射葡萄糖耐量试验IPGTT及其曲线下面积AUCn 6。(H, I) 小鼠腹腔注射胰岛素耐量试验IPITT及其曲线下面积AUCn 6。(J) 小鼠肝脏蛋白质组学数据的KEGG通路富集分析n 4。(K) 普鲁士蓝Perls’ Prussian blue染色显示肝组织铁沉积的代表性图像n 6。(L) 小鼠肝脏铁含量定量分析n 8。(M) 肝脏氧化型磷脂酰胆碱OxPAPC水平n 8。(N) 基于HPLC-MS/MS氧化脂质组学检测的肝脏组织中氧化磷脂OxPLs物种相对丰度变化热图n 5。(O) 差异表达基因左图HFD/Rg3 vs. HFD与差异表达蛋白右图HFD/Rg3 vs. HFD火山图。(P) 肝组织中SPOP蛋白的代表性Western blot图及定量分析。(Q) Rg3与SPOP蛋白的微量热泳动MST结合测定结果。(R) BODIPY 581/591染色的AML12细胞中脂质过氧化水平的代表性共聚焦显微镜图像及定量分析n 3。(S) 流式细胞术检测AML12细胞中线粒体超氧阴离子MitoSOX水平n 3。(T) 示意图Rg3通过上调Spop表达抑制肝细胞脂质过氧化诱导的铁死的作用机制模型。数据以均值 ± 标准误SEM表示*p 0.05**p 0.01。SPOP介导Rg3对AML12细胞铁死亡的保护作用多组学分析一致表明我们采用OxPAPC处理AML12肝细胞建立体外模型。OxPAPC暴露抑制AML12细胞中SPOP的表达而Rg3共处理可有效逆转这一效应图S3A、B。OxPAPC诱导的细胞死亡可被铁死亡抑制剂Ferrostatin-1Fer-1挽救图S3C证实铁死亡是重要的死亡机制。微量热泳动技术MST显示Rg3与SPOP具有高亲和力结合Kd 12.5 nM分子对接模拟鉴定了关键相互作用残基图1Q、S3D-F提示Rg3与SPOP可能存在直接结合。功能上沉默Spop在很大程度上消除了Rg3对OxPLs诱导的细胞死亡的保护作用图S3G。BODIPY 581/591 C11染色显示Spop敲低显著削弱了Rg3介导的脂质过氧化抑制图1R。Rg3可恢复OxPAPC暴露后细胞内GSH水平、GSH/GSSG比值、SOD和CAT活性同时减少MDA积累而这些抗氧化效应在Spop沉默后均受损图S4A-E。此外Spop敲低阻止了Rg3诱导的抗铁死亡酶GPX4的上调图S4F并消除了Rg3介导的OxPAPC处理肝细胞中NADPH水平的恢复图S4G。进一步研究发现Spop缺失消除了Rg3对多种线粒体参数的保护作用图1S、S4H-J。综上这些发现表明SPOP既是Rg3的潜在直接分子靶点也是其在肝细胞中抗铁死亡活性关键的介导因子图1T。SPOP通过促进ATF3泛素化降解抑制铁死亡为鉴定SPOP的下游效应因子我们对AML12细胞中SPOP相互作用蛋白进行了免疫沉淀-质谱IP-MS分析。KEGG分析显示铁死亡相关通路显著富集整合IP-MS结果与体内肝脏蛋白质组学数据鉴定出ATF3是SPOP相互作用蛋白图2A、B。虽然ATF3已被证实与肝脏脂肪变性和葡萄糖代谢紊乱相关但其在代谢性肝病中的作用仍存在争议。我们的结果显示Rg3处理减少了OxPAPC诱导的ATF3蛋白积累图2C且ATF3敲低可部分恢复OxPAPC处理后的GPX4表达图S5A。我们通过GST pull-down实验、Co-IP和分子对接分析证实了SPOP与ATF3之间的直接相互作用Rg3处理显著增加了细胞中SPOP与其底物ATF3的结合量图2D、E、S5B、C。重要的是SPOP通过K48位泛素化促进ATF3的蛋白酶体依赖性降解图2F-I、S6A、B。功能上SPOP过表达可有效抵消AML12细胞中ATF3驱动的铁死亡反应图S6C-F。图2. Rg3通过Spop/Atf3/Nupr1轴缓解肝细胞铁死亡(A)免疫沉淀-质谱IP-MS的KEGG通路富集分析。(B) Venn图中部展示IP-MS鉴定的差异表达蛋白左侧与肝脏蛋白质组学差异蛋白右侧的交集其中ATF3被标记为关键SPOP互作候选蛋白。(C) AML12细胞经溶剂对照、OxPAPC或OxPAPC Rg3处理后ATF3蛋白表达的代表性Western blot及定量结果。(D) 分子对接模拟结果预测SPOP–ATF3蛋白复合物结构及其关键结合残基。(E) GST pull-down实验证实GST标签SPOP与His标签ATF3之间存在直接相互作用。(F) Western blot检测OxPAPC24 h处理下添加溶剂对照、氯喹CQ10 μM自噬抑制剂或MG13210 μM蛋白酶体抑制剂时AML12细胞中ATF3蛋白的稳定性。(G) ATF3共免疫沉淀Co-IP后Western blot检测泛素化修饰证实AML12细胞中ATF3发生泛素化。(H) 在过表达Spop并共转染HA标签泛素质粒的细胞中以抗ATF3抗体进行IP后用抗HA连接特异性抗体进行WB检测。(I) 示意图SPOP介导ATF3的K48连接型泛素化修饰及随后的蛋白酶体降解过程。(J) ChIP-seq代表性峰图显示OxPAPC单独处理EG或联合Rg3处理TG的AML12细胞中ATF3在Nupr1基因位点的结合情况。(K) 双荧光素酶报告基因实验在293T细胞中共转染Atf3表达质粒或空载体对照检测Nupr1启动子活性相对荧光素酶活性以Renilla荧光素酶为内参标准化n 3。(L) 电泳迁移率变动实验EMSA验证ATF3蛋白可直接结合Nupr1启动子区域。(M) ATF3介导Nupr1转录抑制的分子机制模型。(N) 实验设计示意图在CCL4诱导的肝纤维化小鼠模型中采用肝特异性AAV8-shSpop敲低Spop及AAV8-Nupr1过表达Nupr1并给予Rg3干预。(O) CCL₄处理小鼠肝脏中还原型谷胱甘肽GSH水平n 6。(P) CCL4模型小鼠肝脏铁含量n 6。(Q) CCL4模型小鼠肝脏OxPAPC水平n 6。(R) BODIPY 581/591 C11染色的CCL4模型小鼠原代肝细胞中脂质过氧化水平的流式细胞术分析n 3。(S) Seahorse XF分析仪检测CCL4模型小鼠原代肝细胞线粒体生物能量学特征基础及序贯加入寡霉素、FCCP、鱼藤酮/抗霉素A后的耗氧率OCR左图以及最大呼吸能力量化右图n 3。(T) CCL₄模型小鼠肝组织切片中谷胱甘肽过氧化物酶4GPX4表达的代表性免疫荧光图像及定量n 6。(U) 总结模型图Rg3通过Spop–Atf3–Nupr1轴抑制铁死亡从而发挥抗NASH保护作用的分子机制全景图。数据以均值 ± SEM表示p 0.05*p 0.01.Nupr1是ATF3调控肝细胞铁死亡的关键靶点为阐明ATF3调控的转录程序我们对OxPAPC处理及OxPAPC联合Rg3处理的AML12细胞进行了ATF3染色质免疫沉淀测序ChIP-seq图S7、S8A。ATF3结合基因在铁死亡和氧化应激相关通路中显著富集图S8B、C。通过将ChIP-seq数据与通路注释相交我们筛选出4个与氧化应激和铁死亡相关的ATF3结合基因且Atf3敲低可恢复Nupr1和Slc7a11的基因和蛋白表达图2J、S8D、9。鉴于已有研究表明ATF3可调控Slc7a11且Nupr1作为铁死亡的关键抑制因子和氧化还原稳态的调控因子我们将Nupr1作为候选下游效应因子。双荧光素酶报告基因实验和电泳迁移率变动分析EMSA证实ATF3可作为Nupr1的转录抑制因子图2K-M。值得注意的是对肝硬化和铁过载患者肝脏组织的分析显示与正常对照组相比ATF3表达升高SPOP和NUPR1表达下调图S10。Nupr1过表达改善Spop敲低导致的Rg3抗铁死亡功能受损为确定Nupr1是否在Spop下游介导Rg3依赖的铁死亡抑制作用我们在AML12和BRL3A肝细胞中探究了其功能。结果显示Nupr1的重新表达挽救了Spop敲低诱导的Rg3抗氧化效应丧失图S11A-J。类似地Spop敲低显著削弱了Rg3减轻OxPAPC诱导的线粒体损伤的能力而Nupr1过表达在很大程度上逆转了这一效应图S11K、L。Seahorse能量代谢分析显示Rg3恢复了OxPAPC抑制的耗氧率OCR这些效应被Spop敲低消除且在Nupr1过表达后显著恢复图S11M、N。在蛋白水平上Spop沉默减弱了Rg3介导的促铁死亡酶ACSL4的抑制和抗铁死亡蛋白GPX4的诱导而Nupr1可部分改善图S11O、P。Nupr1过表达改善小鼠肝脏中Spop敲低导致的Rg3抗铁死亡效应削弱为在体内验证Nupr1在Spop下游的功能作用我们通过腺病毒递送实现肝脏特异性Spop敲低和Nupr1过表达随后使用四氯化碳CCl4建立肝纤维化模型并通过尾静脉注射OxPAPC诱导肝细胞铁死亡图2N、S12A、B、13A。Spop敲低显著减弱了Rg3的肝脏保护作用包括增强抗氧化能力和减少铁沉积而Nupr1过表达可显著恢复这些作用图2O、P、S12C-E、13B-E、14A-D。类似地沉默Spop抑制了Rg3限制肝脏OxPAPC积累的能力而过表达Nupr1可有效挽救这一效应图2Q、S12F、13F、14C。流式细胞术分析进一步证实Nupr1过表达逆转了Spop敲低导致的Rg3介导的肝脏脂质过氧化抑制丧失图2R、S13G。OCR检测显示Nupr1减轻了Spop敲低诱导的线粒体损伤图2S、S12G、H、13H。在蛋白水平上Spop敲低减少了Rg3介导的ACSL4和F4/80抑制以及GPX4诱导而这些效应在Nupr1过表达后部分恢复图2T、S12I-K、13I、14E、F。结 论本研究系统阐明了Rg3通过抑制OxPLs积累和肝细胞铁死亡缓解NASH的分子机制揭示了Spop-Atf3-Nupr1轴在这一过程中的关键调控作用。Rg3通过激活SPOP促进ATF3泛素化降解解除对Nupr1的转录抑制恢复线粒体稳态减少脂质过氧化最终抑制铁死亡。临床样本分析显示OxPLs水平在NASH患者中显著升高可作为潜在无创生物标志物。这些发现为NASH的发病机制提供了新的见解证实Rg3是靶向Spop-Atf3-Nupr1轴的潜在治疗药物同时为NASH的诊断和治疗提供了新的策略。方 法动物实验设计第一组动物实验图1A6周龄雄性C57BL/6J小鼠饲养在无特定病原体条件下温度控制在22-24℃12小时光照/黑暗周期。小鼠随机分为四组对照组CON喂食标准饲料并每日灌胃溶剂0.5%羧甲基纤维素CMC高脂饮食组HFD喂食高脂饮食60%千卡来自脂肪Research Diets, D1249224周诱导肝脏脂肪变性和纤维化后期每日额外灌胃溶剂HFD低剂量Rg3组HFD/Rg3-L喂食上述高脂饮食并后期每日额外灌胃Rg320 mg/kg体重/天6周HFD高剂量Rg3组HFD/Rg3-H喂食上述高脂饮食并后期每日额外灌胃Rg340 mg/kg体重/天6周。第二组动物实验图2N、S13A在整个实验期间通过胃内灌胃给予小鼠Rg340 mg/kg体重/天。通过单次尾静脉注射编码靶向SPOP的shRNA的AAV8血清型AAV8-shSPOP每只小鼠1×10¹¹病毒基因组vgshRNA序列见表S1实现肝脏特异性SPOP敲低SPOP KD。在Rg3治疗第6周通过单次尾静脉注射AAV8-NUPR1每只小鼠1×10¹¹ vg诱导肝脏NUPR1过表达。AAV8-shSPOP和AAV8-NUPR1均由上海吉玛有限公司设计和生产。为诱导肝铁死亡和肝纤维化建立OxPAPC和CCL4诱导模型。对于OxPAPC模型小鼠每两天尾静脉注射一次OxPAPC4 mg/kg体重持续3周。对于纤维化模型每周三次向小鼠腹腔注射10%v/vCCL4橄榄油剂量为每10 g体重50 μL持续4周。实验结束时处死小鼠收集肝脏组织和血液样本用于后续分析。所有动物实验程序均经浙江省农业科学院动物伦理委员会批准24ZALAS21。人类样本人类肝脏组织样本来自中南大学湘雅医院。所有涉及人类样本的程序均经湘雅医院伦理审查委员会批准人类组织样本伦理批准号2025040650。20(S)-人参皂苷Rg3Rg3的制备与给药Rg3纯度98%成都曼思特生物科技有限公司在给药前溶解于0.5% CMC溶剂中。按指定剂量每日通过胃内灌胃给药6周。对于体外实验Rg3溶解于二甲基亚砜中制备高浓度储备液。细胞培养AML12细胞在添加10%胎牛血清FBS、胰岛素-转铁蛋白-硒、地塞米松和抗生素的DMEM/F12培养基中在37℃、5% CO₂条件下培养。为分离原代小鼠肝细胞麻醉小鼠后先用含EGTA的缓冲液原位灌流肝脏然后用IV型胶原酶消化。将消化后的肝脏切碎通过细胞筛70-100 μm过滤离心50×g5分钟纯化肝细胞。细胞重悬于添加10% FBS、胰岛素、地塞米松、青霉素/链霉素和L-谷氨酰胺的Williams’E培养基中在37℃、5% CO₂条件下培养。接种后4-6小时更换培养基以去除非贴壁细胞。生化检测使用自动生化分析仪测定小鼠血浆中丙氨酸转氨酶ALT、天冬氨酸转氨酶AST、胆固醇TC、甘油三酯TG、乳酸脱氢酶LDH、非酯化脂肪酸NEFA、高密度脂蛋白胆固醇HDL-C和低密度脂蛋白胆固醇LDL-C水平。按照ELISA试剂盒88-7013-2288-7324-2288-7064-22Invitrogen美国马萨诸塞州沃尔瑟姆说明书测定小鼠血浆和肝脏中IL-1β、IL-10和TNFα水平。使用ELISA试剂盒LM-OxLDL-Mu上海联迈生物科技有限公司按照制造商说明书测定血浆OxLDL水平。苏木精-伊红HE和油红OORO染色新鲜肝脏组织样本在室温下用10%福尔马林固定24小时。切成4-5 μm厚的切片并进行HE染色。染色后将切片脱水、透明并封片用于观察。对于ORO染色将冷冻肝脏切片浸入ORO染色液中10-15分钟然后用苏木精复染。随后将切片脱水、透明并封片。使用显微镜Olympus BX41观察脂滴。免疫荧光IF和免疫组织化学IHC染色对于IF染色将石蜡包埋的组织切片脱蜡和水化然后在柠檬酸盐缓冲液中进行抗原修复。切片用0.1% Triton X-100透化用5% BSA封闭然后在4℃下与针对GPX4、ACSL4和F4/80的一抗孵育过夜。洗涤后切片在黑暗中与Alexa Fluor偶联的二抗孵育1小时并用DAPI复染细胞核。对于IHC染色脱蜡和水化后的切片进行抗原修复和内源性过氧化物酶封闭然后在4℃下与针对4-HNE的一抗孵育过夜。随后将切片与HRP偶联的二抗孵育。用DAB底物显色苏木精复染。用显微镜Olympus BX41拍摄图像并通过ImageJ软件定量阳性染色。一抗信息见表S1。透射电子显微镜将新鲜小鼠肝脏组织切成1 mm³的小块或通过离心收集细胞样本用2.5%戊二醛固定4小时然后用1%四氧化锇后固定1-2小时。样本用梯度乙醇脱水包埋在环氧树脂中聚合。使用超薄切片机制备超薄切片~70 nm安装在铜网上用醋酸铀和柠檬酸铅染色。使用透射电子显微镜检查切片并对超微结构特征进行成像和分析。线粒体DNA拷贝数测定使用DNA提取试剂盒货号4403319Thermo Fisher Scientific按照制造商方案从小鼠肝脏组织中分离总DNA。通过定量聚合酶链反应qPCR用线粒体基因组特异性引物定量线粒体DNAmtDNA含量。蛋白质印迹使用含蛋白酶抑制剂的RIPA裂解缓冲液裂解细胞或组织以提取总蛋白收集上清液用于蛋白质浓度测定。提取的蛋白质通过SDS-PAGE分离然后转移到PVDF膜上。膜在4℃下与针对指定靶蛋白的特异性一抗孵育过夜然后在室温下与HRP偶联的二抗孵育2小时。使用成像系统可视化蛋白条带。一抗信息见表S1。微量热泳动对于微量热泳动技术MST实验使用Monolith NT.115仪器NanoTemper Technologies测量平衡解离常数Kd值。简要地使用HIS标签红色通道MO-L018对SPOP蛋白进行荧光标记将Rg3稀释至所需浓度范围从200 μM到0.0061 μM并与0.1 μM荧光标记的SPOP蛋白孵育15分钟。将样本加载到NanoTemper玻璃毛细管MO-K025中在40%发光二极管功率下进行微量热泳动。通过NanoTemper软件使用质量作用方程从重复读数计算Kd值。每个实验至少进行三次。免疫共沉淀Co-IP和GST pull-down对于Co-IP用RIPA缓冲液裂解细胞以提取总蛋白。将澄清的裂解液在4℃下与指定的一抗或对照IgG孵育过夜以形成免疫复合物。然后加入Protein A/G琼脂糖珠将混合物再孵育2小时以捕获抗体-蛋白复合物。用洗涤缓冲液洗涤珠子以去除非特异性结合的蛋白质。最后洗脱结合的蛋白质并通过SDS-PAGE分离通过蛋白质印迹分析。所有实验中均包含IgG对照以评估非特异性结合。对于GST-pull down实验将纯化的GST-SPOP融合蛋白固定在谷胱甘肽琼脂糖珠上通过洗涤去除未结合的蛋白质。然后将珠子与含有指定靶蛋白的裂解液孵育以形成蛋白质-蛋白质复合物。经过广泛洗涤去除非特异性相互作用物后使用洗脱缓冲液洗脱结合的蛋白质通过SDS-PAGE分离并通过蛋白质印迹分析。单独的GST蛋白用作阴性对照。细胞线粒体耗氧率OCR将细胞以1×10⁴/孔接种在Seahorse XFe 96孔板中。处理后将培养基更换为不含缓冲液的Seahorse XF培养基pH 7.4含有10 mM葡萄糖、1 mM丙酮酸钠和2 mM GlutaMAX然后在37℃、无CO₂条件下孵育1小时。依次注射化合物终浓度1 μM寡霉素、0.5 μM FCCP、0.5 μM鱼藤酮/抗霉素A以评估基础呼吸、最大呼吸和非线粒体耗氧。使用XFe分析仪记录OCR值并通过Seahorse Wave软件进行分析。流式细胞术分析收集细胞用PBS洗涤并用MitoSOX Red或BODIPY™ 581/591 C11染色37℃30分钟黑暗中。未染色的细胞用作阴性对照。用PBS洗涤两次后重悬细胞并通过流式细胞术分析。使用FlowJo软件定量荧光强度并将数据归一化至阴性对照。双荧光素酶报告基因测定将NUPR1启动子片段亚克隆到pGL3-Basic载体中以生成萤火虫荧光素酶报告基因构建体。将人胚肾293T细胞接种在24孔板中使用Lipofectamine 3000将报告质粒、ATF3过表达质粒或空载体阴性对照与pRL-TK海肾荧光素酶质粒内参共转染。转染48小时后使用双荧光素酶报告基因检测系统测量荧光素酶活性。相对荧光素酶活性计算为萤火虫荧光素酶活性与海肾荧光素酶活性的比值。RNA测序RNA测序由上海欧易生物医学科技有限公司进行。简要地从小鼠肝脏组织中提取总RNA使用Qubit 4.0进行浓度测量使用Agilent Bioanalyzer进行片段大小分布和完整性评估。将测序文库标准化后在Illumina NovaSeq 6000平台上以双端模式PE150测序每个样本生成10-20百万条读数以确保低丰度转录本的覆盖。使用FastQC评估原始数据质量使用Trimmomatic过滤低质量读数/接头。通过HISAT2或STAR将清洁读数比对到参考基因组。使用StringTie或Salmon定量基因/转录本表达水平FPKM或TPM。使用DESeq2或edgeR鉴定差异表达基因通过调整p值或错误发现率FDR 0.05确定显著性。使用clusterProfiler或DAVID对显著基因集进行功能富集分析GO和KEGG通路q值 0.05的通路被认为显著富集。蛋白质组学分析使用数据非依赖采集DIA工作流程进行蛋白质组学分析。简要地从肝脏组织中提取的蛋白质用胰蛋白酶消化、脱盐并通过反相超高效液相色谱UHPLCThermo Vanquish Neo系统使用50 cm Low-Load µPAC™ Neo HPLC柱分离。流动相A0.1%甲酸水溶液和B0.1%甲酸在80%乙腈中以2.2 μL/min的流速进行8分钟线性梯度洗脱。在DIA模式下操作Orbitrap Astral质谱仪进行肽段分析全MS扫描380-980 m/z分辨率240,000AGC500%注射时间5 ms和DIA MS/MS扫描150-2000 m/z隔离窗口2 m/zAGC500%注射时间3 ms碰撞能量为25%循环时间为0.6秒。全MS和DIA光谱分别以轮廓模式和质心模式记录。使用MaxQuant或SpectronautUniProt数据库处理原始数据前体质量公差为±10 ppm胰蛋白酶/P特异性≤2个漏切位点。基于倍数变化≥1.5和调整p值 0.05鉴定差异表达蛋白质。然后使用DAVID和STRING对这些蛋白质进行功能富集分析Benjamini-Hochberg调整p值 0.05的富集术语被认为显著。免疫沉淀-质谱IP-MS分析对于IP-MS分析在OxPAPC暴露前用氯喹和MG132预处理AML12细胞6小时然后提取蛋白质。将细胞裂解液与偶联到Protein A/G琼脂糖珠的抗SPOP抗体孵育以形成抗原-抗体复合物。作为阴性对照使用同型匹配的IgG进行平行免疫沉淀。使用珠子分离免疫复合物并通过广泛洗涤去除非特异性结合的蛋白质。洗脱结合的蛋白质并进行LC-MS/MS分析以进行蛋白质鉴定和定量。使用MaxQuant或类似平台处理数据以确定每种蛋白质的无标记定量LFQ强度。为鉴定高可信度相互作用蛋白质对SPOP-IP和IgG-IP重复样品之间的LFQ强度进行双样本t检验。使用Benjamini-Hochberg方法调整所得p值。如果蛋白质满足FDR 0.05和倍数富集 2的标准则被认为是相互作用蛋白。基于LC-MS/MS的氧化脂质组学分析将组织样本在750 μL氯仿甲醇超纯水3:6:1v/v/v中匀浆在4℃下轻轻振荡1500 rpm孵育1小时。通过加入350 μL去离子水和250 μL氯仿诱导相分离然后离心。将含有脂质的下层有机相收集到干净的管中。为最大化脂质回收率通过向剩余水相中加入450 μL氯仿重复提取一次。合并有机提取物使用SpeedVac在OH模式下真空干燥。脂质组学分析在LipidALL Technologies使用ExionLC-AD—Sciex QTRAP 6500 PLUS进行。通过正相NPHPLC使用TUP-HB硅胶柱内径150 mm×2.1 mm3 μm实现氧化磷脂类别的分离。设置多反应监测MRM转变用于各种氧化磷脂的比较分析。通过参考加标的内标定量各个脂质种类。DMPC和DMPE购自Avanti Polar Lipids。然后使用Benjamini-Hochberg程序调整所得p值。FDR 0.05的脂质被认为显著改变。染色质免疫沉淀测序ChIP-seq分析用1%甲醛固定细胞以交联DNA-蛋白质复合物然后用甘氨酸淬灭。裂解细胞通过超声破碎染色质至平均大小为200-500 bp。将破碎的染色质在4℃下与ATF3特异性抗体和Protein A/G磁珠Thermo Fisher Scientific孵育过夜。平行进行同型匹配的IgG免疫沉淀作为背景对照。洗涤后洗脱复合物并逆转交联。纯化DNA制备测序文库并进行测序。对原始数据进行质量控制、接头修剪并比对到参考基因组mm10Bowtie2。通过将ATF3 ChIP-seq数据与IgG对照数据进行比较使用MACS2进行峰调用并使用FDR阈值 0.05鉴定显著峰。随后对显著峰集进行基序分析MEME Suite和可视化Integrative Genomics ViewerIGV。数据统计分析所有定量数据均以均值 ± 标准误mean ± SEM 表示其中n代表生物学重复数。采用 GraphPad Prism 9 软件美国加利福尼亚州拉霍亚市GraphPad 公司进行统计学分析。两组间比较采用学生氏t检验Studentst-test三组及以上独立样本间的比较适当时采用单因素方差分析one-way ANOVA 并经Tukey 多重比较检验 进行后续分析。对于重复测量数据采用线性混合效应模型LMM 分析模型分析后再通过Tukey 多重比较检验 进行组间配对的事后比较。所有检验中p值0.05 被认为具有统计学显著性。代码和数据可用性本研究所使用的原始测序数据已存储在国家生物信息中心CNCB/BIG数据库中登录号CRA024779 和 CRA024780。质谱蛋白质组学数据已保存至 ProteomeXchange 联盟的 iProX 存储库项目编号IPX001516700密码cpxN。主要数据和代码已上传至 GitHub。组学数据分析通过OE生物云平台https://cloud.oebiotech.com完成。补充材料可从 iMeta Science (http://www.imeta.science/) 获取。引文格式Yibo Zong, Guo Long, Tiantian Gu, Pan Huang, Yong Tian, Wenwu Xu, Xiheng Hu, et al. 2026. “A Bioactive Ginsenoside Alleviates Hepatocellular Ferroptosis Induced by Oxidized Phospholipid Accumulation in Nonalcoholic Steatohepatitis.”iMeta5: e70114. https://doi.org/10.1002/imt2.70114.B1作者简介结 论总体而言1999年以来美国绝经后女性RA死亡率显著下降提示治疗进步与医疗服务改善已带来实质性获益。然而不同种族/族裔、地区及年龄群体之间仍存在显著差异。未来应通过更精准的公共卫生策略与医疗公平干预改善高风险人群的诊疗可及性与健康结局。宗一博第一作者● 浙江省农业科学院畜牧兽医研究所 博后。● 主要研究方向为应激生物学和动物福利脂质代谢和免疫调控。主持国家自然科学基金青年基金参与国家自然科学基金面上项目及国家重点研发计划课题/子课题以第一在iMeta、antioxidants、poultry science等杂志上发表SCI论文5篇。结 论总体而言1999年以来美国绝经后女性RA死亡率显著下降提示治疗进步与医疗服务改善已带来实质性获益。然而不同种族/族裔、地区及年龄群体之间仍存在显著差异。未来应通过更精准的公共卫生策略与医疗公平干预改善高风险人群的诊疗可及性与健康结局。史达尊通讯作者● 中南大学湘雅医院副主任医师硕士生导师中华医学会妇科肿瘤学会青年委员。● 主要研究方向为肿瘤耐药机制及天然产物提取物的临床应用、新型抗肿瘤药物研发等。主持国家博士后科学基金、湖南省自然科学基金等国家及省部级项目2项以第一/通讯作者在Journal of Advanced Research、Gynecologic Oncology、laboratory investigation等行业高影响力杂志上发表原创性SCI论文16篇。结 论总体而言1999年以来美国绝经后女性RA死亡率显著下降提示治疗进步与医疗服务改善已带来实质性获益。然而不同种族/族裔、地区及年龄群体之间仍存在显著差异。未来应通过更精准的公共卫生策略与医疗公平干预改善高风险人群的诊疗可及性与健康结局。卢立志通讯作者● 浙江省农业科学院畜牧兽医研究所研究员博士生导师。● 研究方向为家禽重要性状遗传机理解析、育种新技术研发、新品种培育及产业化配套技术。浙江省特级专家神农领军英才浙江省万人计划杰出人才浙江省农业科技突出贡献者。主持国家重点研发计划、国家自然科学基金浙江省重点研发等国家及省部级科研项目20余项以第一/通讯作者在iMeta、Genome Biology、Journal of Nanobiotechnology等国内外期刊发表学术论文200多篇主持获国家科技进步二等奖1项省科技进步一等奖2项。曾涛通讯作者● 浙江省农业科学院畜牧兽医研究所副所长副研究员博士生导师。● 研究方向为家禽重要性状遗传机制解析、天然提取物生物学功能及在家禽生产应用等研究。主持国家重点研发计划课题/子课题、国家自然科学基金项目面上和青年、浙江省自然科学基金重点项目等国家及省部级项目10余项。以第一/通讯作者在iMeta、Communications Biology、Food Chemistry等期刊发表SCI论文30余篇参与获国家科技进步二等奖1项省科技进步一等奖2项。共同主办单位更多推荐▼ 点击跳转高引文章 ▸▸▸▸iMeta | 引用20000海普洛斯陈实富发布新版fastp更快更好地处理FASTQ数据高引文章 ▸▸▸▸iMeta | 兰大张东组使用PhyloSuite进行分子系统发育及系统发育树的统计分析高引文章▸▸▸▸iMeta | 唐海宝/张兴坦-用于比较基因组学分析的多功能分析套件JCVIiMeta封面1卷1期1卷2期1卷3期1卷4期2卷1期2卷2期2卷3期2卷4期3卷1期3卷2期3卷3期3卷4期3卷5期3卷6期4卷1期4卷2期4卷3期4卷4期4卷5期4卷6期iMetaOmics封面1卷1期1卷2期2卷1期2卷2期2卷3期2卷4期iMetaMed封面1卷1期1卷2期期刊简介“iMeta” 是由威立、宏科学和本领域数千名华人科学家合作出版的开放获取期刊主编由中科院微生物所刘双江研究员和荷兰格罗宁根大学傅静远教授担任。目的是发表所有领域高影响力的研究、方法和综述重点关注微生物组、生物信息、大数据和多组学等前沿交叉学科。目标是发表前10%(IF 20)的高影响力论文。期刊特色包括中英双语图文、双语视频、可重复分析、图片打磨、60万用户的社交媒体宣传等。2022年2月正式创刊相继被Google Scholar、PubMed、SCIE、ESI、DOAJ、Scopus等数据库收录2025年6月影响因子33.2中科院分区生物学1区Top位列全球SCI期刊前千分之三(65/22249)微生物学科2/163仅低于Nature Reviews学科研究类期刊全球第一中国大陆5/585“iMetaOmics” 是“iMeta” 子刊主编由中国科学院北京生命科学研究院赵方庆研究员和香港中文大学于君教授担任目标是成为影响因子大于10的高水平综合期刊欢迎投稿iMetaMed 是“iMeta” 子刊专注于医学、健康和生物技术领域目标是成为影响因子大于15的医学综合类期刊欢迎投稿iMeta主页http://www.imeta.science姊妹刊iMetaOmics主页http://www.imeta.science/imetaomics/出版社iMeta主页https://onlinelibrary.wiley.com/journal/2770596x出版社iMetaOmics主页https://onlinelibrary.wiley.com/journal/29969514出版社iMetaMed主页https://onlinelibrary.wiley.com/journal/3066988xiMeta投稿https://wiley.atyponrex.com/journal/IMT2iMetaOmics投稿https://wiley.atyponrex.com/journal/IMO2iMetaMed投稿https://wiley.atyponrex.com/submission/dashboard?siteNameIMM3邮箱officeimeta.science
本文来自互联网用户投稿,该文观点仅代表作者本人,不代表本站立场。本站仅提供信息存储空间服务,不拥有所有权,不承担相关法律责任。如若转载,请注明出处:http://www.coloradmin.cn/o/2475152.html
如若内容造成侵权/违法违规/事实不符,请联系多彩编程网进行投诉反馈,一经查实,立即删除!相关文章
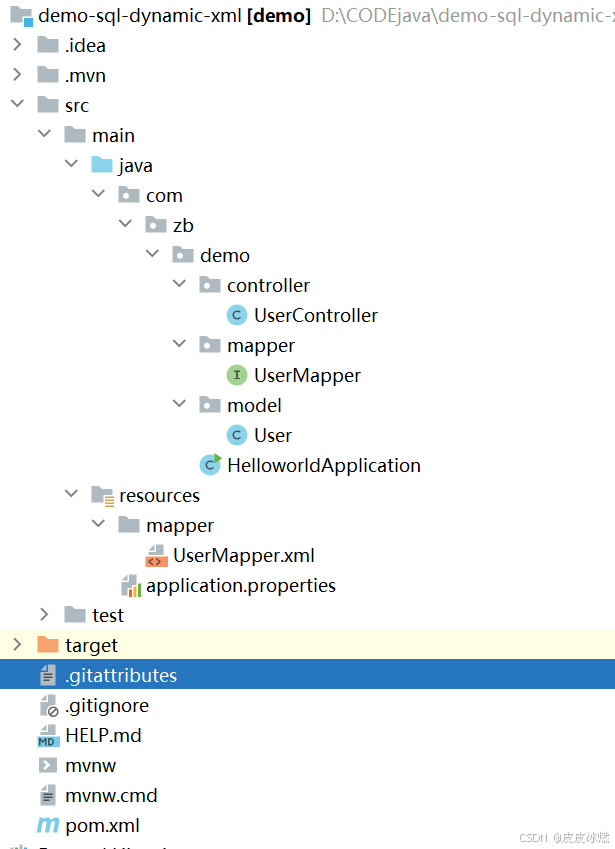
SpringBoot-17-MyBatis动态SQL标签之常用标签
文章目录 1 代码1.1 实体User.java1.2 接口UserMapper.java1.3 映射UserMapper.xml1.3.1 标签if1.3.2 标签if和where1.3.3 标签choose和when和otherwise1.4 UserController.java2 常用动态SQL标签2.1 标签set2.1.1 UserMapper.java2.1.2 UserMapper.xml2.1.3 UserController.ja…
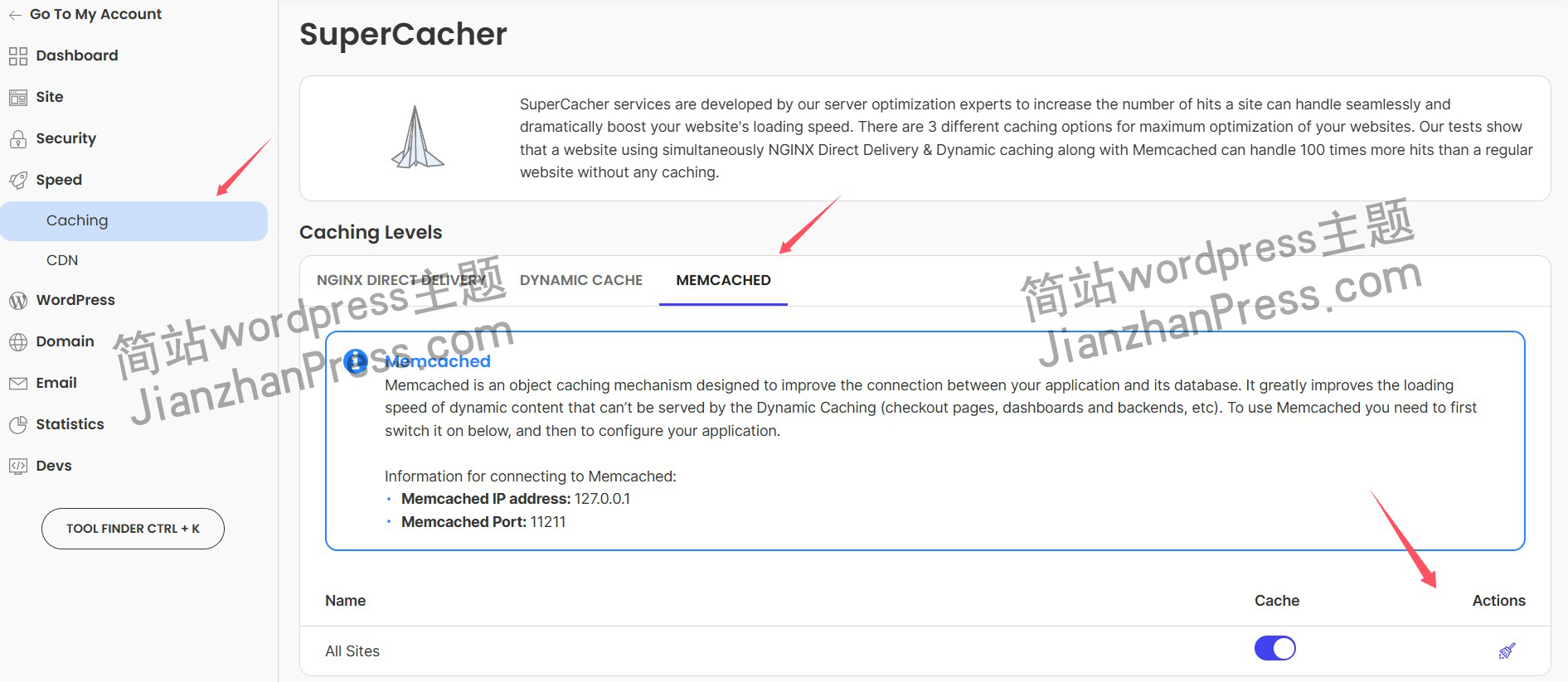
wordpress后台更新后 前端没变化的解决方法
使用siteground主机的wordpress网站,会出现更新了网站内容和修改了php模板文件、js文件、css文件、图片文件后,网站没有变化的情况。
不熟悉siteground主机的新手,遇到这个问题,就很抓狂,明明是哪都没操作错误&#x…
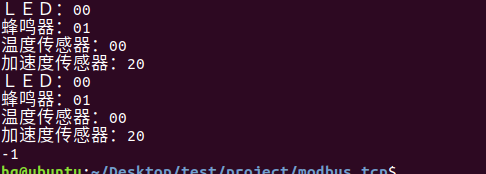
网络编程(Modbus进阶)
思维导图 Modbus RTU(先学一点理论)
概念 Modbus RTU 是工业自动化领域 最广泛应用的串行通信协议,由 Modicon 公司(现施耐德电气)于 1979 年推出。它以 高效率、强健性、易实现的特点成为工业控制系统的通信标准。 包…
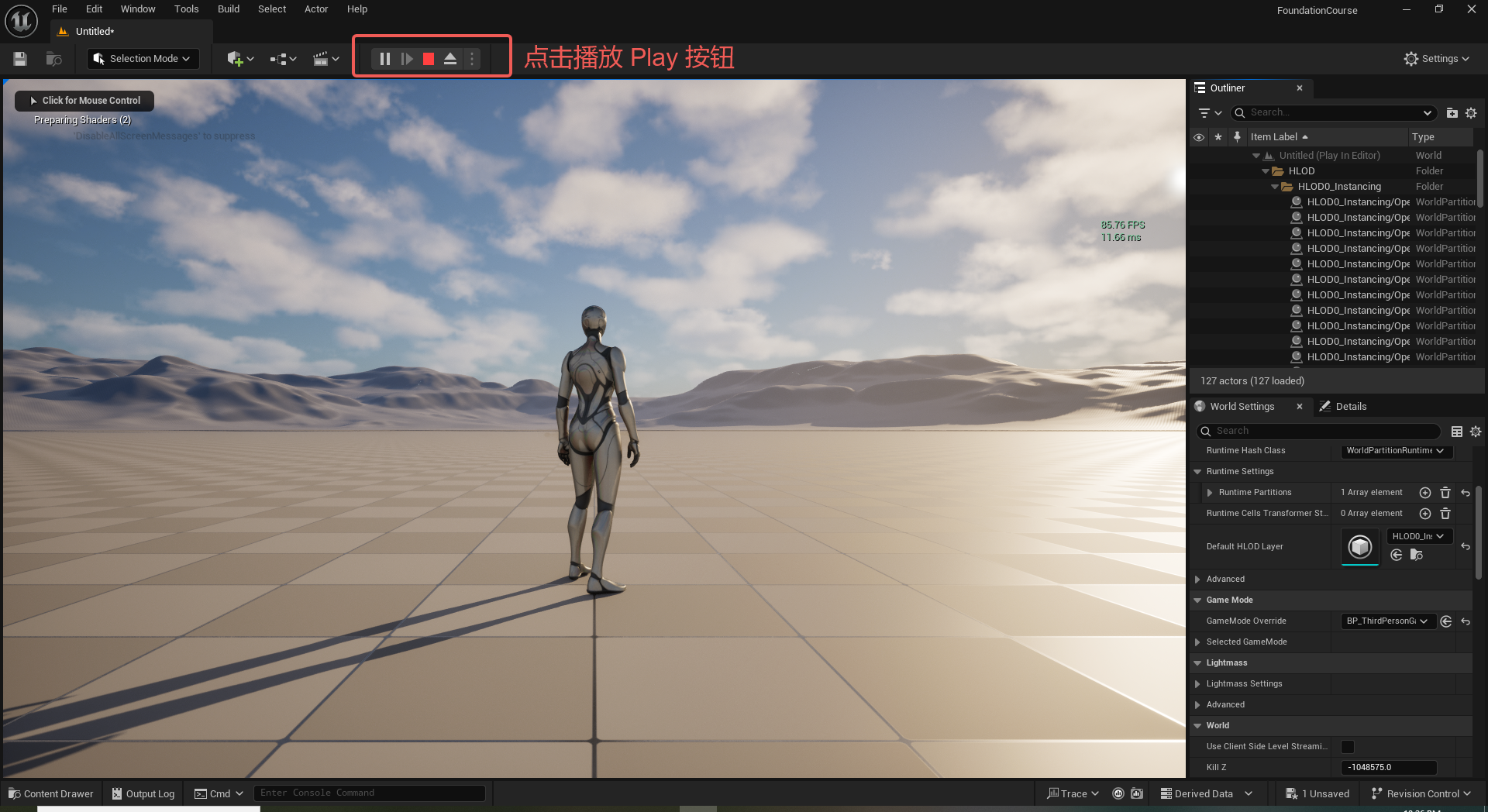
UE5 学习系列(二)用户操作界面及介绍
这篇博客是 UE5 学习系列博客的第二篇,在第一篇的基础上展开这篇内容。博客参考的 B 站视频资料和第一篇的链接如下:
【Note】:如果你已经完成安装等操作,可以只执行第一篇博客中 2. 新建一个空白游戏项目 章节操作,重…
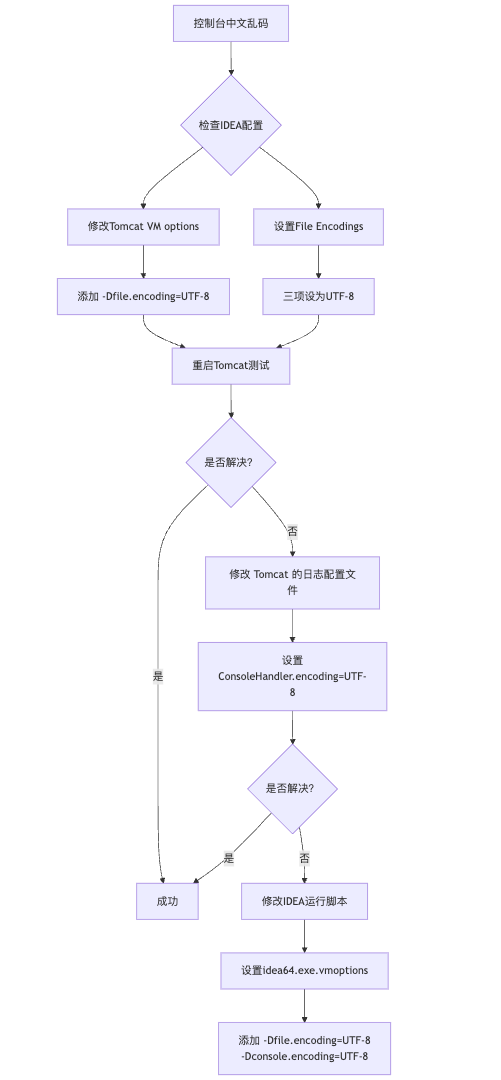
IDEA运行Tomcat出现乱码问题解决汇总
最近正值期末周,有很多同学在写期末Java web作业时,运行tomcat出现乱码问题,经过多次解决与研究,我做了如下整理:
原因:
IDEA本身编码与tomcat的编码与Windows编码不同导致,Windows 系统控制台…

利用最小二乘法找圆心和半径
#include <iostream>
#include <vector>
#include <cmath>
#include <Eigen/Dense> // 需安装Eigen库用于矩阵运算 // 定义点结构
struct Point { double x, y; Point(double x_, double y_) : x(x_), y(y_) {}
}; // 最小二乘法求圆心和半径 …
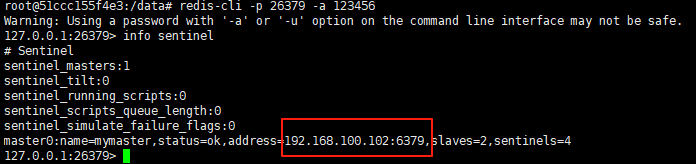
使用docker在3台服务器上搭建基于redis 6.x的一主两从三台均是哨兵模式
一、环境及版本说明
如果服务器已经安装了docker,则忽略此步骤,如果没有安装,则可以按照一下方式安装: 1. 在线安装(有互联网环境): 请看我这篇文章 传送阵>> 点我查看 2. 离线安装(内网环境):请看我这篇文章 传送阵>> 点我查看
说明:假设每台服务器已…
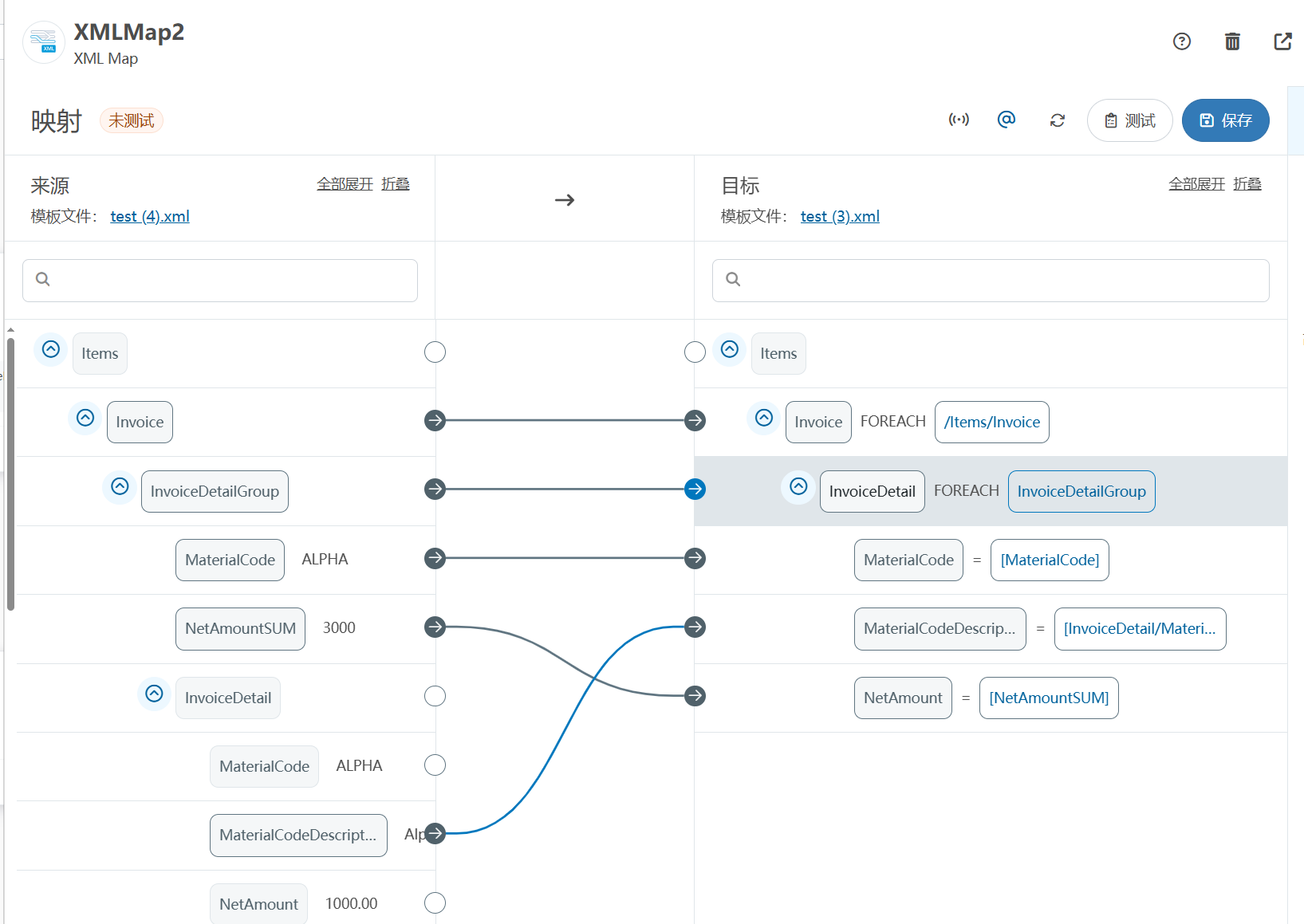
XML Group端口详解
在XML数据映射过程中,经常需要对数据进行分组聚合操作。例如,当处理包含多个物料明细的XML文件时,可能需要将相同物料号的明细归为一组,或对相同物料号的数量进行求和计算。传统实现方式通常需要编写脚本代码,增加了开…
LBE-LEX系列工业语音播放器|预警播报器|喇叭蜂鸣器的上位机配置操作说明
LBE-LEX系列工业语音播放器|预警播报器|喇叭蜂鸣器专为工业环境精心打造,完美适配AGV和无人叉车。同时,集成以太网与语音合成技术,为各类高级系统(如MES、调度系统、库位管理、立库等)提供高效便捷的语音交互体验。
L…

(LeetCode 每日一题) 3442. 奇偶频次间的最大差值 I (哈希、字符串)
题目:3442. 奇偶频次间的最大差值 I 思路 :哈希,时间复杂度0(n)。 用哈希表来记录每个字符串中字符的分布情况,哈希表这里用数组即可实现。
C版本:
class Solution {
public:int maxDifference(string s) {int a[26]…

【大模型RAG】拍照搜题技术架构速览:三层管道、两级检索、兜底大模型
摘要
拍照搜题系统采用“三层管道(多模态 OCR → 语义检索 → 答案渲染)、两级检索(倒排 BM25 向量 HNSW)并以大语言模型兜底”的整体框架: 多模态 OCR 层 将题目图片经过超分、去噪、倾斜校正后,分别用…
【Axure高保真原型】引导弹窗
今天和大家中分享引导弹窗的原型模板,载入页面后,会显示引导弹窗,适用于引导用户使用页面,点击完成后,会显示下一个引导弹窗,直至最后一个引导弹窗完成后进入首页。具体效果可以点击下方视频观看或打开下方…
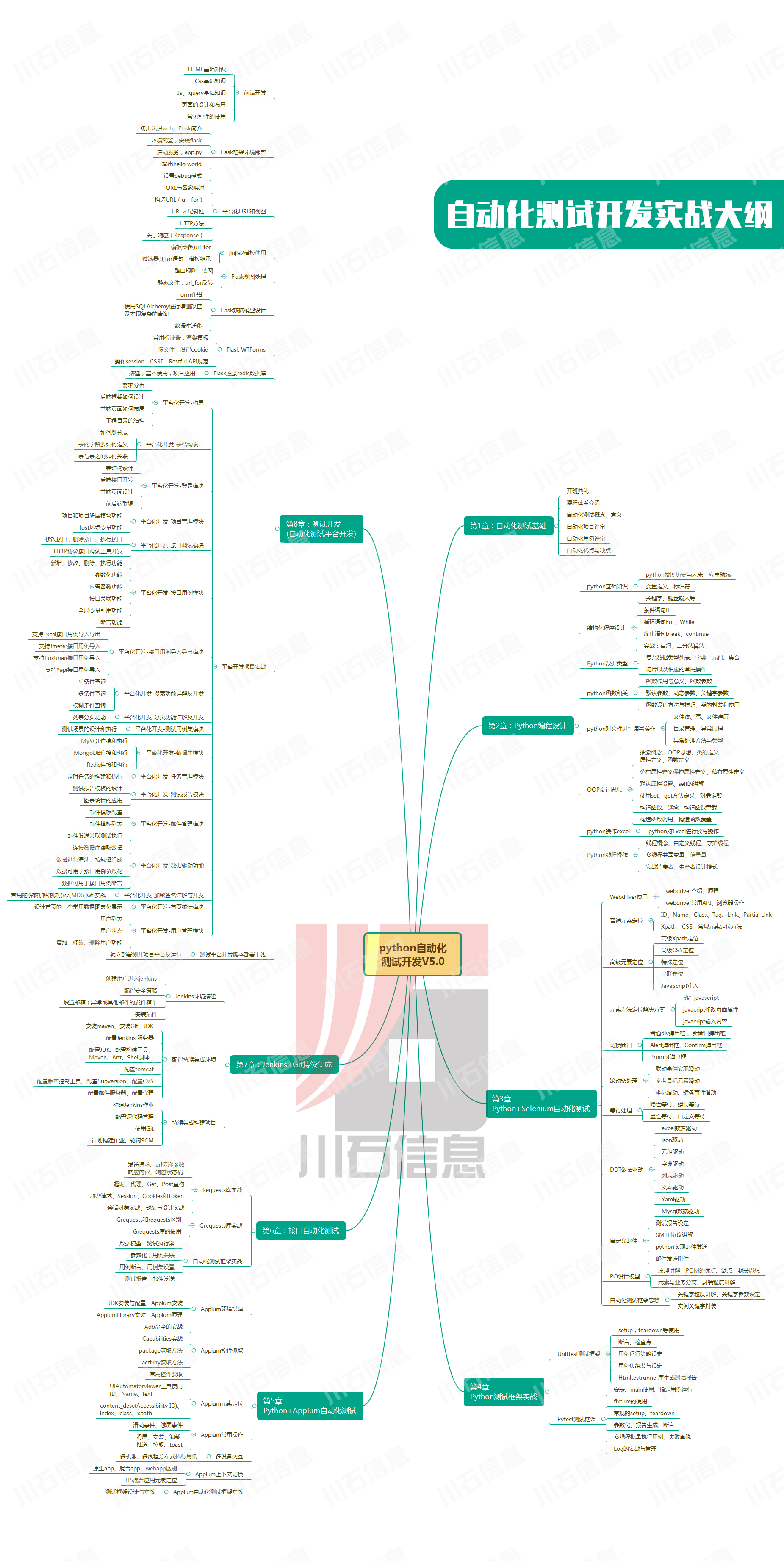
接口测试中缓存处理策略
在接口测试中,缓存处理策略是一个关键环节,直接影响测试结果的准确性和可靠性。合理的缓存处理策略能够确保测试环境的一致性,避免因缓存数据导致的测试偏差。以下是接口测试中常见的缓存处理策略及其详细说明:
一、缓存处理的核…
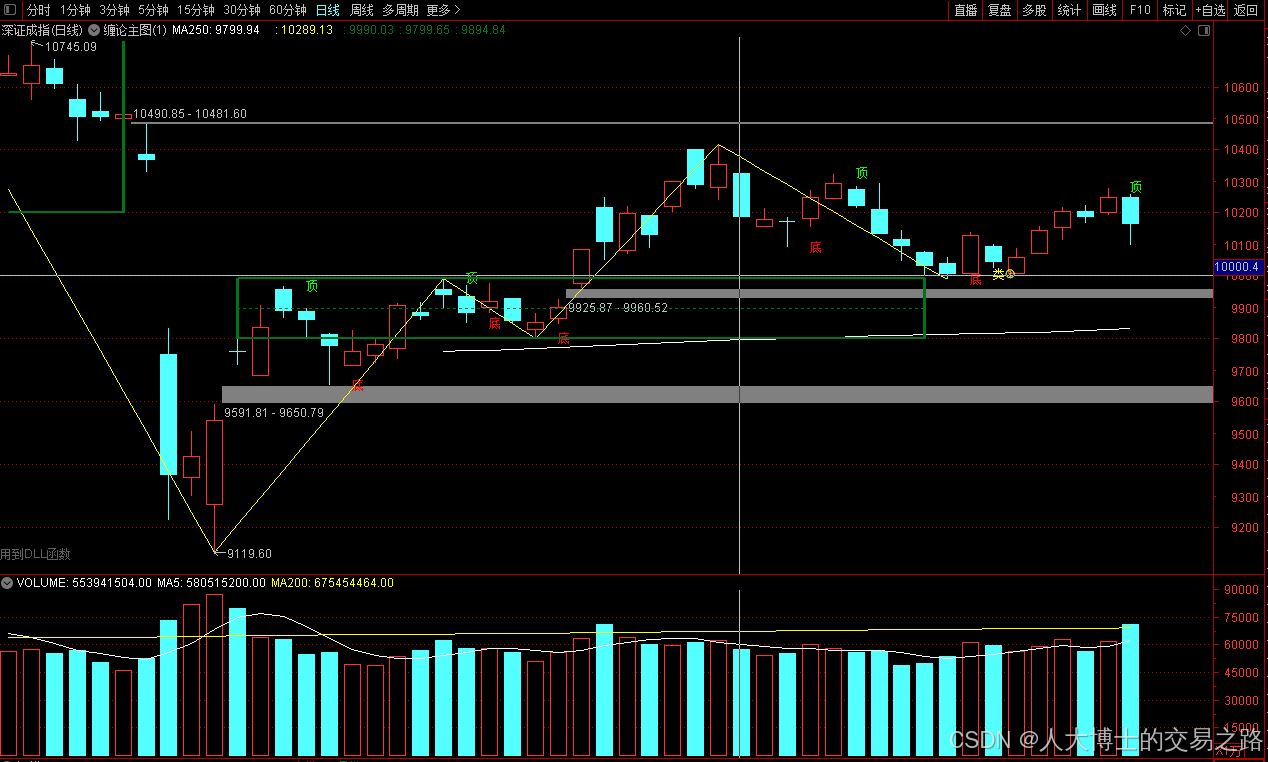
龙虎榜——20250610
上证指数放量收阴线,个股多数下跌,盘中受消息影响大幅波动。 深证指数放量收阴线形成顶分型,指数短线有调整的需求,大概需要一两天。 2025年6月10日龙虎榜行业方向分析 1. 金融科技
代表标的:御银股份、雄帝科技
驱动…
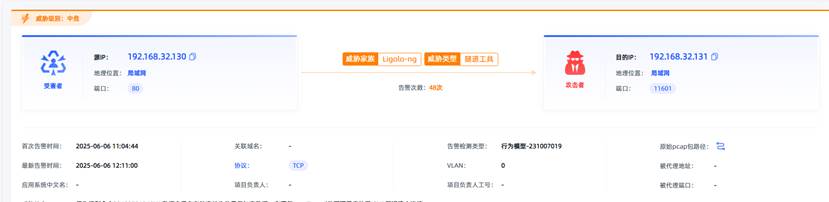
观成科技:隐蔽隧道工具Ligolo-ng加密流量分析
1.工具介绍
Ligolo-ng是一款由go编写的高效隧道工具,该工具基于TUN接口实现其功能,利用反向TCP/TLS连接建立一条隐蔽的通信信道,支持使用Let’s Encrypt自动生成证书。Ligolo-ng的通信隐蔽性体现在其支持多种连接方式,适应复杂网…

铭豹扩展坞 USB转网口 突然无法识别解决方法
当 USB 转网口扩展坞在一台笔记本上无法识别,但在其他电脑上正常工作时,问题通常出在笔记本自身或其与扩展坞的兼容性上。以下是系统化的定位思路和排查步骤,帮助你快速找到故障原因:
背景:
一个M-pard(铭豹)扩展坞的网卡突然无法识别了,扩展出来的三个USB接口正常。…

未来机器人的大脑:如何用神经网络模拟器实现更智能的决策?
编辑:陈萍萍的公主一点人工一点智能 未来机器人的大脑:如何用神经网络模拟器实现更智能的决策?RWM通过双自回归机制有效解决了复合误差、部分可观测性和随机动力学等关键挑战,在不依赖领域特定归纳偏见的条件下实现了卓越的预测准…

Linux应用开发之网络套接字编程(实例篇)
服务端与客户端单连接
服务端代码
#include <sys/socket.h>
#include <sys/types.h>
#include <netinet/in.h>
#include <stdio.h>
#include <stdlib.h>
#include <string.h>
#include <arpa/inet.h>
#include <pthread.h>
…

华为云AI开发平台ModelArts
华为云ModelArts:重塑AI开发流程的“智能引擎”与“创新加速器”!
在人工智能浪潮席卷全球的2025年,企业拥抱AI的意愿空前高涨,但技术门槛高、流程复杂、资源投入巨大的现实,却让许多创新构想止步于实验室。数据科学家…

深度学习在微纳光子学中的应用
深度学习在微纳光子学中的主要应用方向
深度学习与微纳光子学的结合主要集中在以下几个方向:
逆向设计 通过神经网络快速预测微纳结构的光学响应,替代传统耗时的数值模拟方法。例如设计超表面、光子晶体等结构。
特征提取与优化 从复杂的光学数据中自…