单细胞转录组分析流程:从细胞矩阵生成到聚类、注释与轨迹推断
news2026/4/1 16:12:31
点击“AladdinEdu你的AI学习实践工作坊”注册即送-H卡级别算力沉浸式云原生集成开发环境80G大显存多卡并行按量弹性计费教育用户更享超低价。摘要单细胞RNA测序scRNA-seq技术的普及使得在单细胞分辨率下解析转录组异质性成为可能但海量数据的分析对生物信息学方法提出了巨大挑战。本文系统阐述scRNA-seq数据分析的完整流程从原始数据预处理、表达矩阵生成、质量控制、标准化、特征选择、降维可视化到细胞聚类、差异表达分析、细胞类型注释及轨迹推断。深入解析每个步骤的核心算法原理如PCA、t-SNE、UMAP、Leiden聚类、RNA速率等和主流工具Seurat、Scanpy、Monocle的使用方法并结合实际案例演示如何构建可靠的分析管线。通过本文读者可掌握从单细胞测序数据到生物学发现的全流程分析技能。关键词单细胞转录组scRNA-seq细胞聚类细胞注释轨迹推断Seurat1. 引言单细胞RNA测序single-cell RNA sequencing, scRNA-seq技术自2009年诞生以来已彻底改变了我们对细胞异质性的理解。与传统的bulk RNA-seq不同scRNA-seq能够在单细胞分辨率下解析转录组揭示稀有细胞类型、发育轨迹、细胞状态转换以及细胞间相互作用。然而scRNA-seq数据的分析远比bulk RNA-seq复杂数据具有高维度数万个基因×数千至数十万个细胞、高稀疏性大量零值、高噪声技术变异和生物学变异混杂等特点需要专门的分析方法和流程。经过十余年的发展scRNA-seq数据分析已形成一套相对标准化的流程包括原始数据处理、质量控制、标准化、特征选择、降维、聚类、差异表达、细胞注释和轨迹推断等步骤。本文将以最常用的R包Seurat和Python库Scanpy为主线系统介绍每个步骤的原理、方法和实践技巧帮助研究者构建可靠的分析管线。2. 数据预处理与表达矩阵生成2.1 原始数据处理scRNA-seq的原始数据通常以FASTQ格式存储需要经过预处理才能得到细胞-基因表达矩阵。预处理步骤包括读段比对使用STAR、STARsolo、Cell Ranger等工具将读段比对到参考基因组。细胞barcode识别根据barcode序列将读段分配到特定细胞微滴平台。UMI去重利用UMIUnique Molecular Identifier去除PCR扩增重复实现绝对定量。基因计数统计每个细胞中每个基因的UMI数量生成表达矩阵。主流处理工具10x Genomics Cell Ranger10x平台数据的标准处理流程输出矩阵文件.h5或mtx格式。STARsoloSTAR比对器的单细胞模式支持多种平台。Drop-seq ToolsDrop-seq数据的处理工具。kallisto | bustools轻量级定量方法适用于多种平台。输出格式表达矩阵通常以稀疏矩阵matrix.mtx存储同时包含基因列表features.tsv和细胞barcode列表barcodes.tsv。2.2 读入表达矩阵到分析环境SeuratRlibrary(Seurat)# 读入10x数据data_dir-path/to/filtered_feature_bc_matrix/counts-Read10X(data_dir)seurat_obj-CreateSeuratObject(countscounts,projectscRNA_project,min.cells3,min.features200)ScanpyPythonimportscanpyassc adatasc.read_10x_mtx(path/to/filtered_feature_bc_matrix/,gex_onlyTrue)adata.var_names_make_unique()3. 质量控制scRNA-seq数据中包含大量低质量细胞如死细胞、双胞体和低表达基因需要在分析前进行过滤。3.1 细胞水平质控指标nCount_RNA每个细胞检测到的UMI总数。过低可能表示细胞已死亡或文库构建失败过高可能表示双胞体。nFeature_RNA每个细胞检测到的基因数。与nCount相关过低表示低质量细胞。线粒体基因比例percent.mt线粒体基因表达比例过高通常10-20%提示细胞应激或凋亡。核糖体基因比例percent.rb某些情况下也可用于质控。Seurat质控示例# 计算线粒体基因比例seurat_obj[[percent.mt]]-PercentageFeatureSet(seurat_obj,pattern^MT-)# 可视化VlnPlot(seurat_obj,featuresc(nFeature_RNA,nCount_RNA,percent.mt),ncol3)# 过滤seurat_obj-subset(seurat_obj,subsetnFeature_RNA200nFeature_RNA5000percent.mt15)Scanpy质控示例adata.var[mt]adata.var_names.str.startswith(MT-)sc.pp.calculate_qc_metrics(adata,qc_vars[mt],percent_topNone,log1pFalse,inplaceTrue)sc.pl.violin(adata,[n_genes_by_counts,total_counts,pct_counts_mt],jitter0.4,multi_panelTrue)adataadata[adata.obs.n_genes_by_counts5000,:]adataadata[adata.obs.pct_counts_mt15,:]3.2 基因水平过滤移除在极少细胞中表达的基因如3个细胞可降低数据稀疏性减少计算负担。Seuratseurat_obj-subset(seurat_obj,featuresrownames(seurat_obj)[Matrix::rowSums(GetAssayData(seurat_obj)0)3])3.3 双胞体检测双胞体doublet指一个微滴中包含两个细胞会产生异常的基因表达模式干扰聚类结果。常用检测方法DoubletFinder基于人工合成双胞体计算每个细胞的双胞体得分。ScrubletPython类似原理。Seurat DoubletFinderlibrary(DoubletFinder)sweep_res-paramSweep_v3(seurat_obj,PCs1:20,sctFALSE)sweep_stats-summarizeSweep(sweep_res,GTFALSE)bcmvn-find.pK(sweep_stats)pK-as.numeric(as.character(bcmvn$pK[bcmvn$BCmetricmax(bcmvn$BCmetric)]))seurat_obj-doubletFinder_v3(seurat_obj,PCs1:20,pN0.25,pKpK,nExpncol(seurat_obj)*0.04)# 过滤双胞体seurat_obj-subset(seurat_obj,cellsrownames(seurat_objmeta.data)[seurat_objmeta.data$DF.classificationsSinglet])4. 标准化由于不同细胞的测序深度不同需要标准化以消除技术差异使表达水平具有可比性。4.1 对数标准化最常用的方法是将每个细胞的UMI计数除以该细胞的总计数乘以缩放因子通常为10,000再取对数log1p。公式log( (counts / total_counts) * 10000 1 )Seuratseurat_obj-NormalizeData(seurat_obj,normalization.methodLogNormalize,scale.factor10000)4.2 SCTransformSeurat提供的SCTransform方法利用正则化负二项回归模型同时标准化和去除技术变异包括测序深度和线粒体基因比例效果优于传统LogNormalize尤其适用于批次效应校正前的预处理。seurat_obj-SCTransform(seurat_obj,vars.to.regresspercent.mt)Scanpysc.pp.normalize_total(adata,target_sum1e4)sc.pp.log1p(adata)5. 特征选择数据集中大多数基因在不同细胞间表达变化较小对区分细胞类型贡献有限。特征选择旨在筛选出表达变异性高的基因用于后续降维和聚类。5.1 高变基因HVG识别常用方法计算每个基因的均值和方差拟合均值-方差关系选择方差显著高于预期的基因。Seurat# 方法1: 传统FindVariableFeaturesseurat_obj-FindVariableFeatures(seurat_obj,selection.methodvst,nfeatures2000)# 方法2: SCTransform后自动识别seurat_obj-SCTransform(seurat_obj,variable.features.n3000)Scanpysc.pp.highly_variable_genes(adata,n_top_genes2000,flavorseurat_v3)adata.rawadata adataadata[:,adata.var.highly_variable]6. 降维可视化高维数据难以直接可视化需要降维到2维或3维空间。6.1 主成分分析PCAPCA是最常用的线性降维方法将原始特征投影到方差最大的正交轴上。PCA结果用于后续聚类和轨迹推断。Seurat# 在SCTransform后自动计算PCAseurat_obj-RunPCA(seurat_obj,npcs50)ElbowPlot(seurat_obj)# 确定选择的主成分数通常选择拐点处或之前6.2 t-SNE与UMAPt-SNE非线性降维擅长保持局部结构但无法保持全局距离计算速度慢随机性较大。UMAP非线性降维同时保持局部和部分全局结构速度更快是目前单细胞可视化的主流选择。Seuratseurat_obj-RunUMAP(seurat_obj,dims1:30)# 使用前30个PCDimPlot(seurat_obj,reductionumap)Scanpysc.pp.neighbors(adata,n_neighbors30,n_pcs30)sc.tl.umap(adata)sc.pl.umap(adata,colorsample)7. 聚类聚类将具有相似表达谱的细胞归为一群用于识别细胞类型或状态。7.1 基于图的聚类单细胞聚类普遍采用基于共享最近邻图SNN的方法构建细胞-细胞相似性图基于PCA空间。优化图结构如SNN。使用社区发现算法如Louvain、Leiden识别聚类。Seurat# 构建SNN图并聚类seurat_obj-FindNeighbors(seurat_obj,dims1:30)seurat_obj-FindClusters(seurat_obj,resolution0.8)# resolution控制聚类粒度DimPlot(seurat_obj,labelTRUE)Scanpysc.pp.neighbors(adata,n_neighbors15,n_pcs30)sc.tl.leiden(adata,resolution0.8)sc.pl.umap(adata,colorleiden)7.2 聚类分辨率选择低分辨率0.1-0.5聚类数少适合识别主要细胞类型。高分辨率0.8-2.0聚类数多适合识别精细亚群。通常需要结合生物学知识和可视化结果进行调整。8. 差异表达分析与细胞类型注释8.1 差异表达分析识别每个聚类相对于其他聚类的标记基因用于推断细胞类型。SeuratFindAllMarkers# 默认使用Wilcoxon秩和检验markers-FindAllMarkers(seurat_obj,only.posTRUE,min.pct0.25,logfc.threshold0.25)# 按聚类查看top标记基因top_markers-markers%%group_by(cluster)%%top_n(5,avg_log2FC)DoHeatmap(seurat_obj,featurestop_markers$gene)Scanpyrank_genes_groupssc.tl.rank_genes_groups(adata,groupbyleiden,methodwilcoxon)sc.pl.rank_genes_groups(adata,n_genes5,shareyFalse)8.2 细胞类型注释细胞注释是将聚类与已知细胞类型对应起来。常用方法人工注释基于已知标记基因结合文献和数据库如CellMarker、PanglaoDB。自动注释使用参考数据集如SingleR、scmap、CellAssign进行标签转移。Seurat SingleR需结合R包library(SingleR)# 准备参考数据集如BlueprintEncodeDataref-celldex::BlueprintEncodeData()# 进行注释seurat_obj$singleR_labels-SingleR(testGetAssayData(seurat_obj,slotdata),refref,labelsref$label.main)9. 批次效应校正当数据来自多个批次不同样本、不同测序日期、不同平台时批次效应可能混淆生物学差异需要进行校正。9.1 常用方法Harmony将数据投影到共享的嵌入空间迭代校正批次效应。CCA典型相关分析Seurat的IntegrateData方法基于CCA。BBKNNScanpy中的批次校正方法。Scanorama适用于大规模数据。Seurat Harmonylibrary(harmony)seurat_obj-RunHarmony(seurat_obj,group.by.varsbatch,reductionpca,dims.use1:30)seurat_obj-RunUMAP(seurat_obj,reductionharmony,dims1:30)seurat_obj-FindNeighbors(seurat_obj,reductionharmony,dims1:30)Scanpy BBKNNsc.external.pp.bbknn(adata,batch_keybatch,neighbors_within_batch3)sc.tl.umap(adata)10. 轨迹推断当细胞处于连续的分化或激活状态时聚类无法反映细胞间的关系需要轨迹推断trajectory inference来重建细胞状态转变路径。10.1 常用方法Monocle基于逆向图嵌入构建拟时序轨迹。Slingshot基于聚类结果的曲线推断支持多条轨迹。PAGA基于图的抽象分析可视化细胞群间的连接。RNA速率RNA velocity利用未剪接和剪接mRNA的比例推断分化方向。10.2 Monocle 3Seurat集成library(SeuratWrappers)library(monocle3)cds-as.cell_data_set(seurat_obj)cds-cluster_cells(cds,reduction_methodUMAP)cds-learn_graph(cds)cds-order_cells(cds)plot_cells(cds,color_cells_bypseudotime)10.3 SlingshotRlibrary(slingshot)sce-as.SingleCellExperiment(seurat_obj)sce-slingshot(sce,clusterLabelsseurat_clusters,reducedDimUMAP)10.4 RNA速率Python, scVeloimportscveloasscv adatascv.read(adata_with_unspliced.h5ad)scv.pp.filter_and_normalize(adata)scv.pp.moments(adata)scv.tl.velocity(adata)scv.tl.velocity_graph(adata)scv.pl.velocity_embedding_stream(adata,basisumap)11. 实战案例外周血单核细胞PBMC分析11.1 数据下载使用10x Genomics提供的3k PBMC数据集# 下载数据library(Seurat)pbmc_data-Read10X(path/to/pbmc3k_filtered_gene_bc_matrices/hg19/)pbmc-CreateSeuratObject(countspbmc_data,projectpbmc3k,min.cells3,min.features200)11.2 完整分析流程# 质控pbmc[[percent.mt]]-PercentageFeatureSet(pbmc,pattern^MT-)pbmc-subset(pbmc,subsetnFeature_RNA200nFeature_RNA2500percent.mt5)# 标准化与特征选择pbmc-NormalizeData(pbmc)pbmc-FindVariableFeatures(pbmc,nfeatures2000)# 降维pbmc-ScaleData(pbmc)pbmc-RunPCA(pbmc)ElbowPlot(pbmc)pbmc-RunUMAP(pbmc,dims1:10)# 聚类pbmc-FindNeighbors(pbmc,dims1:10)pbmc-FindClusters(pbmc,resolution0.5)DimPlot(pbmc,labelTRUE)# 标记基因与注释pbmc.markers-FindAllMarkers(pbmc,only.posTRUE,min.pct0.25,logfc.threshold0.25)# 根据已知标记注释CD3D (T细胞), CD14 (单核细胞), MS4A1 (B细胞), etc.new.cluster.ids-c(CD4 T细胞,CD14单核细胞,B细胞,CD8 T细胞,NK细胞,CD16单核细胞,DC细胞,血小板)names(new.cluster.ids)-levels(pbmc)pbmc-RenameIdents(pbmc,new.cluster.ids)DimPlot(pbmc,labelTRUE)12. 挑战与未来方向12.1 当前挑战数据稀疏性大量dropout事件导致真实信号丢失。批次效应多批次数据整合仍需改进。细胞注释标准化缺乏统一的细胞类型命名体系。大规模数据计算百万级细胞数据对内存和计算资源要求极高。空间信息丢失单细胞测序破坏空间结构。12.2 未来趋势多组学整合同时分析转录组、表观组、蛋白质组。空间转录组结合空间位置信息解析组织微环境。深度学习应用使用变分自编码器VAE进行批次校正、插补和细胞状态建模。动态分析从静态快照推断动态过程的方法不断改进。13. 结语单细胞转录组数据分析是一个复杂但高度结构化的流程从原始数据处理到生物学发现每一步都有成熟的算法和工具支持。Seurat和Scanpy作为两大主流分析框架提供了完整的分析功能从基础的质控、标准化到高级的轨迹推断和RNA速率分析。掌握这些工具的核心原理和操作是开展单细胞研究的必备技能。随着技术的快速发展单细胞分析正从单一模态向多模态、从静态向动态、从细胞悬液向空间原位演进。未来的数据分析方法将继续创新以应对更复杂的数据结构和更深层次的生物学问题。参考文献Stuart, T., et al. (2019). Comprehensive integration of single-cell data.Cell, 177(7), 1888-1902.Wolf, F. A., et al. (2018). SCANPY: large-scale single-cell gene expression data analysis.Genome Biology, 19(1), 15.Korsunsky, I., et al. (2019). Fast, sensitive and accurate integration of single-cell data with Harmony.Nature Methods, 16(12), 1289-1296.Cao, J., et al. (2019). The single-cell transcriptional landscape of mammalian organogenesis.Nature, 566(7745), 496-502.La Manno, G., et al. (2018). RNA velocity of single cells.Nature, 560(7719), 494-498.点击“AladdinEdu你的AI学习实践工作坊”注册即送-H卡级别算力沉浸式云原生集成开发环境80G大显存多卡并行按量弹性计费教育用户更享超低价。
本文来自互联网用户投稿,该文观点仅代表作者本人,不代表本站立场。本站仅提供信息存储空间服务,不拥有所有权,不承担相关法律责任。如若转载,请注明出处:http://www.coloradmin.cn/o/2438373.html
如若内容造成侵权/违法违规/事实不符,请联系多彩编程网进行投诉反馈,一经查实,立即删除!相关文章
SpringBoot-17-MyBatis动态SQL标签之常用标签
文章目录 1 代码1.1 实体User.java1.2 接口UserMapper.java1.3 映射UserMapper.xml1.3.1 标签if1.3.2 标签if和where1.3.3 标签choose和when和otherwise1.4 UserController.java2 常用动态SQL标签2.1 标签set2.1.1 UserMapper.java2.1.2 UserMapper.xml2.1.3 UserController.ja…
wordpress后台更新后 前端没变化的解决方法
使用siteground主机的wordpress网站,会出现更新了网站内容和修改了php模板文件、js文件、css文件、图片文件后,网站没有变化的情况。
不熟悉siteground主机的新手,遇到这个问题,就很抓狂,明明是哪都没操作错误&#x…
网络编程(Modbus进阶)
思维导图 Modbus RTU(先学一点理论)
概念 Modbus RTU 是工业自动化领域 最广泛应用的串行通信协议,由 Modicon 公司(现施耐德电气)于 1979 年推出。它以 高效率、强健性、易实现的特点成为工业控制系统的通信标准。 包…
UE5 学习系列(二)用户操作界面及介绍
这篇博客是 UE5 学习系列博客的第二篇,在第一篇的基础上展开这篇内容。博客参考的 B 站视频资料和第一篇的链接如下:
【Note】:如果你已经完成安装等操作,可以只执行第一篇博客中 2. 新建一个空白游戏项目 章节操作,重…
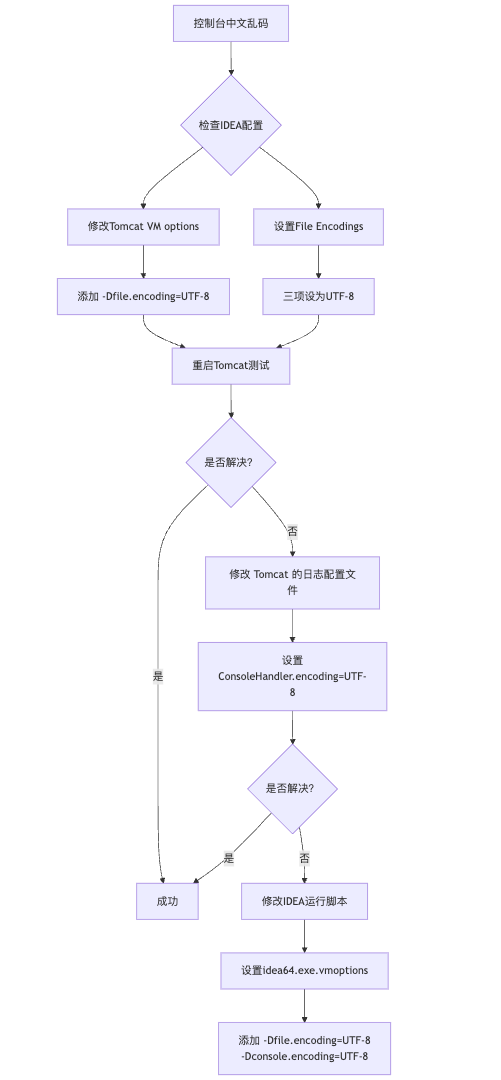
IDEA运行Tomcat出现乱码问题解决汇总
最近正值期末周,有很多同学在写期末Java web作业时,运行tomcat出现乱码问题,经过多次解决与研究,我做了如下整理:
原因:
IDEA本身编码与tomcat的编码与Windows编码不同导致,Windows 系统控制台…

利用最小二乘法找圆心和半径
#include <iostream>
#include <vector>
#include <cmath>
#include <Eigen/Dense> // 需安装Eigen库用于矩阵运算 // 定义点结构
struct Point { double x, y; Point(double x_, double y_) : x(x_), y(y_) {}
}; // 最小二乘法求圆心和半径 …
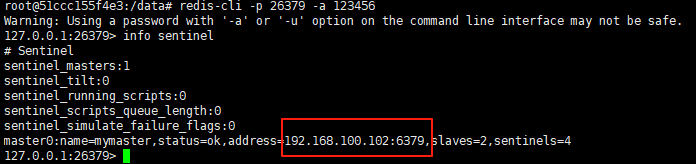
使用docker在3台服务器上搭建基于redis 6.x的一主两从三台均是哨兵模式
一、环境及版本说明
如果服务器已经安装了docker,则忽略此步骤,如果没有安装,则可以按照一下方式安装: 1. 在线安装(有互联网环境): 请看我这篇文章 传送阵>> 点我查看 2. 离线安装(内网环境):请看我这篇文章 传送阵>> 点我查看
说明:假设每台服务器已…
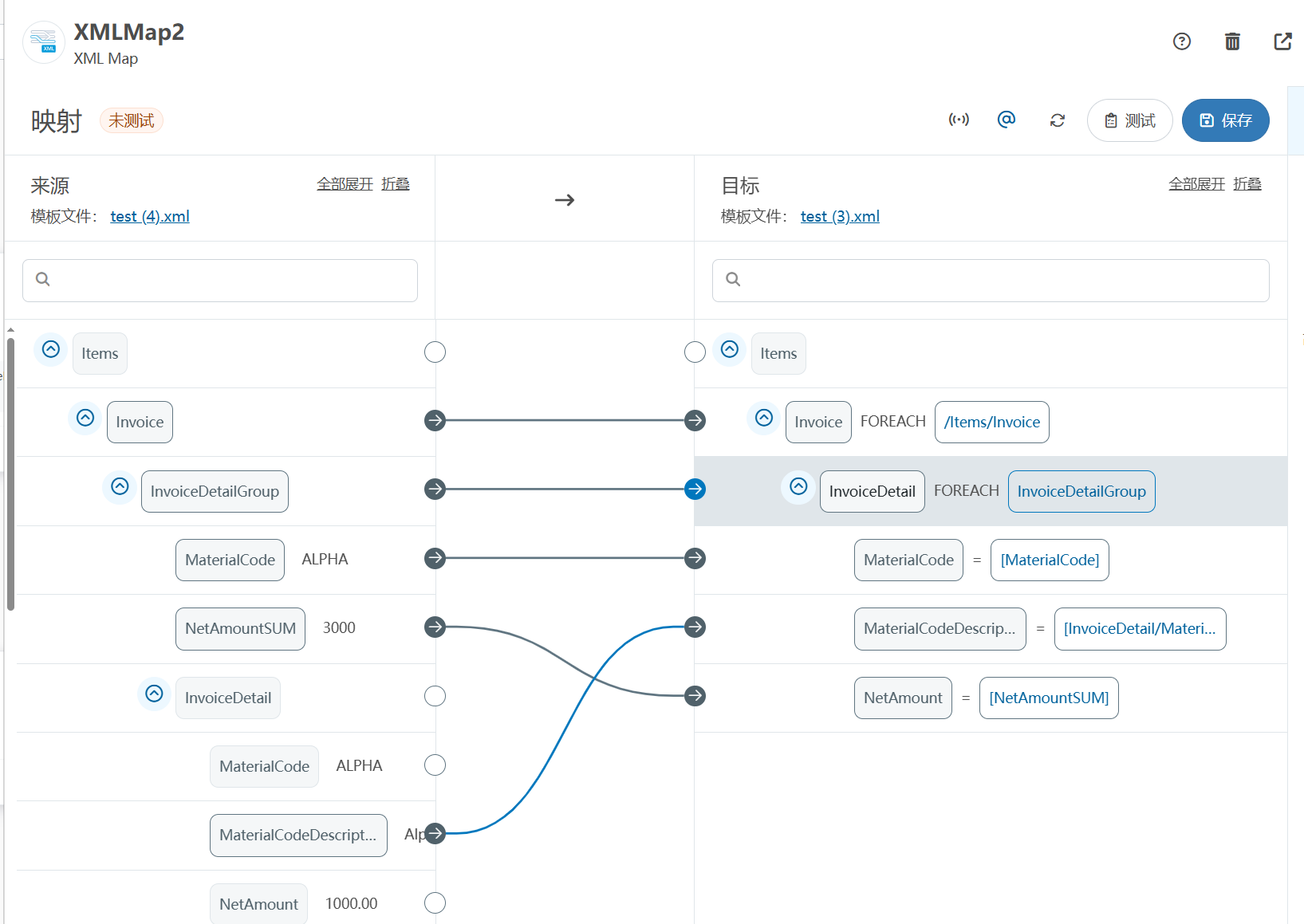
XML Group端口详解
在XML数据映射过程中,经常需要对数据进行分组聚合操作。例如,当处理包含多个物料明细的XML文件时,可能需要将相同物料号的明细归为一组,或对相同物料号的数量进行求和计算。传统实现方式通常需要编写脚本代码,增加了开…
LBE-LEX系列工业语音播放器|预警播报器|喇叭蜂鸣器的上位机配置操作说明
LBE-LEX系列工业语音播放器|预警播报器|喇叭蜂鸣器专为工业环境精心打造,完美适配AGV和无人叉车。同时,集成以太网与语音合成技术,为各类高级系统(如MES、调度系统、库位管理、立库等)提供高效便捷的语音交互体验。
L…

(LeetCode 每日一题) 3442. 奇偶频次间的最大差值 I (哈希、字符串)
题目:3442. 奇偶频次间的最大差值 I 思路 :哈希,时间复杂度0(n)。 用哈希表来记录每个字符串中字符的分布情况,哈希表这里用数组即可实现。
C版本:
class Solution {
public:int maxDifference(string s) {int a[26]…

【大模型RAG】拍照搜题技术架构速览:三层管道、两级检索、兜底大模型
摘要
拍照搜题系统采用“三层管道(多模态 OCR → 语义检索 → 答案渲染)、两级检索(倒排 BM25 向量 HNSW)并以大语言模型兜底”的整体框架: 多模态 OCR 层 将题目图片经过超分、去噪、倾斜校正后,分别用…
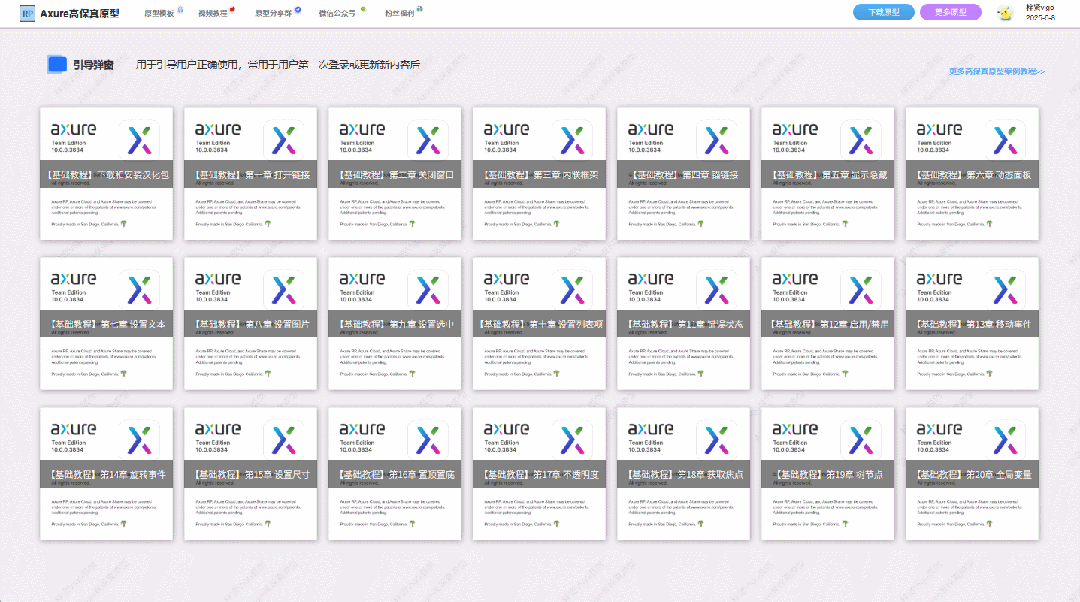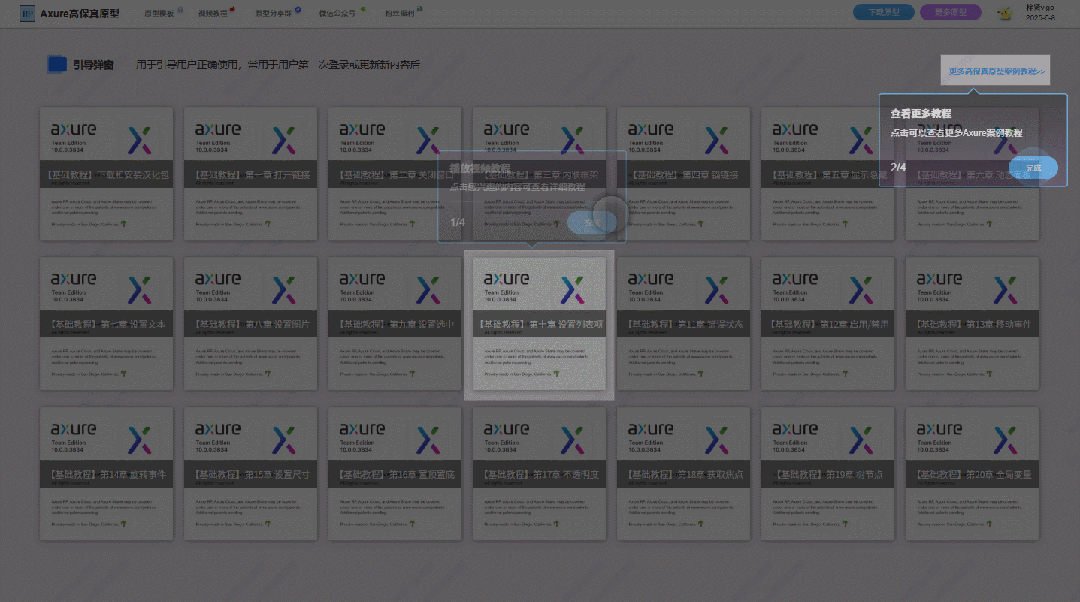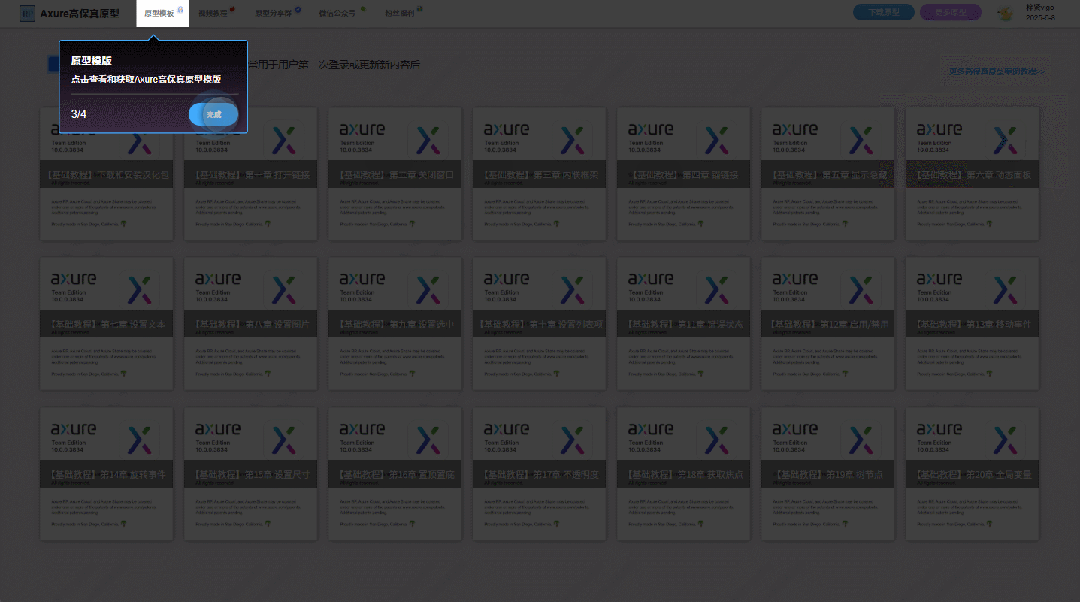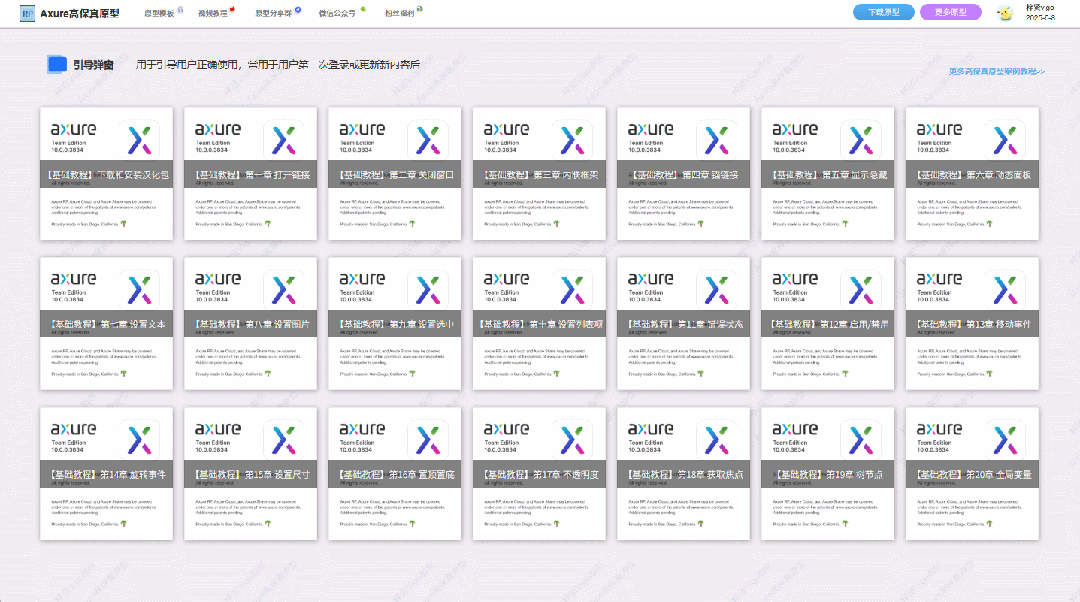
【Axure高保真原型】引导弹窗
今天和大家中分享引导弹窗的原型模板,载入页面后,会显示引导弹窗,适用于引导用户使用页面,点击完成后,会显示下一个引导弹窗,直至最后一个引导弹窗完成后进入首页。具体效果可以点击下方视频观看或打开下方…
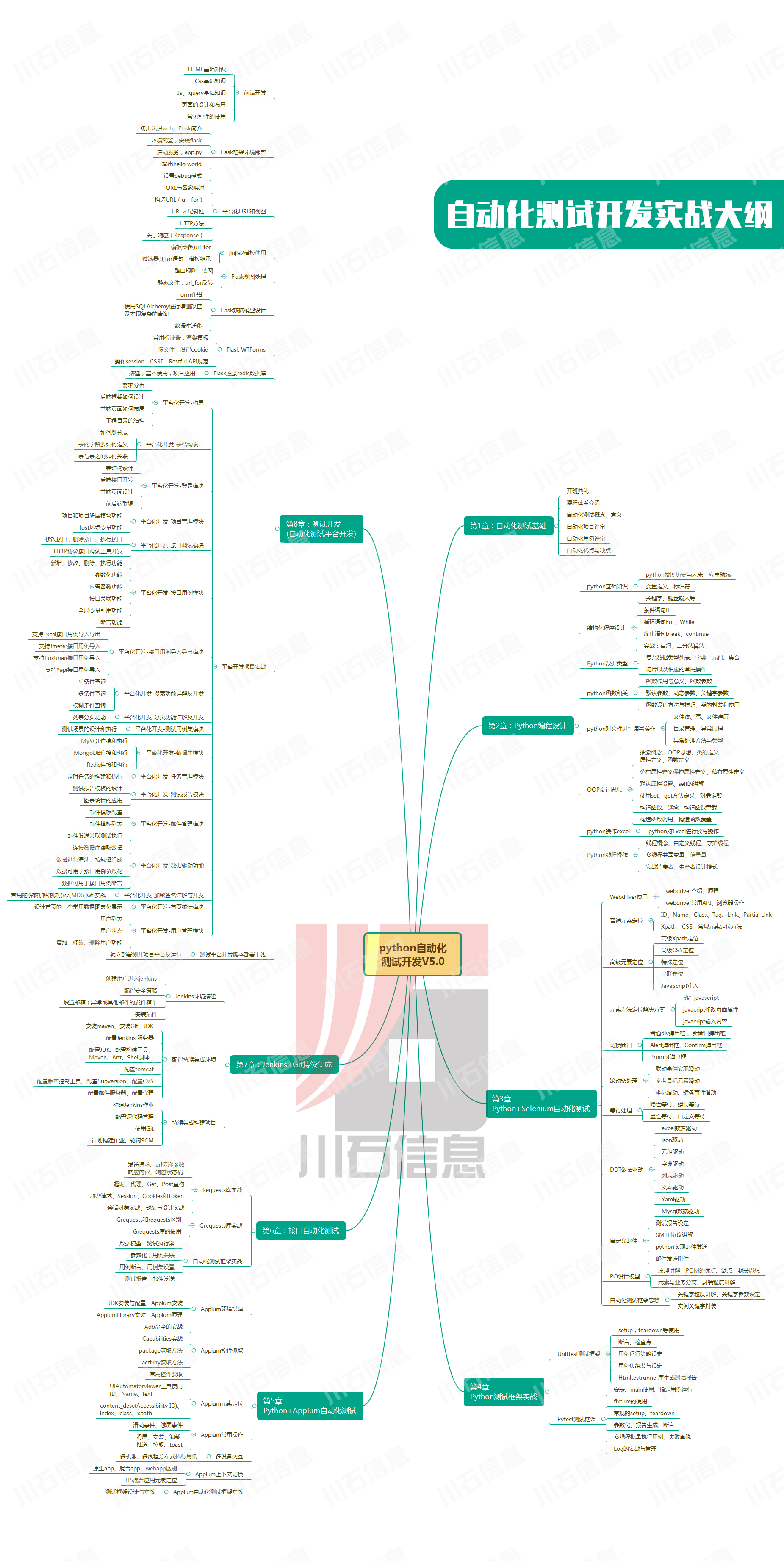
接口测试中缓存处理策略
在接口测试中,缓存处理策略是一个关键环节,直接影响测试结果的准确性和可靠性。合理的缓存处理策略能够确保测试环境的一致性,避免因缓存数据导致的测试偏差。以下是接口测试中常见的缓存处理策略及其详细说明:
一、缓存处理的核…
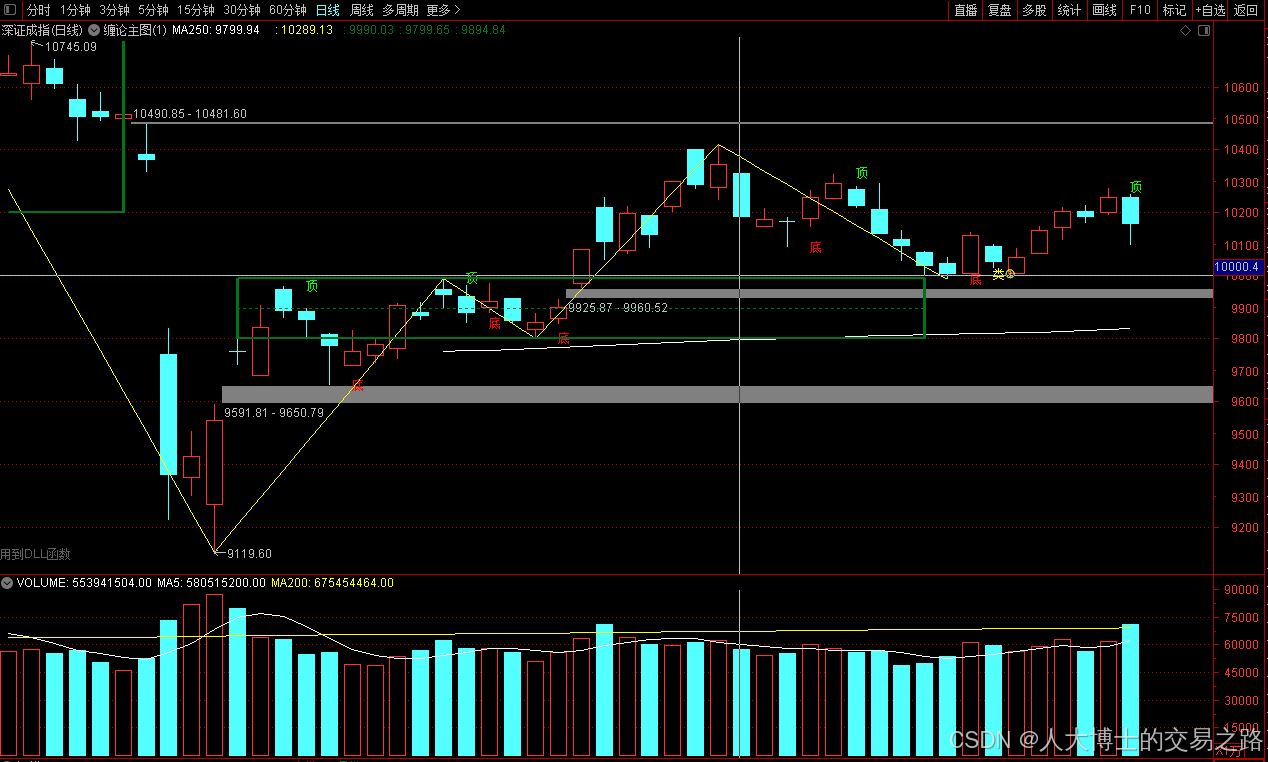
龙虎榜——20250610
上证指数放量收阴线,个股多数下跌,盘中受消息影响大幅波动。 深证指数放量收阴线形成顶分型,指数短线有调整的需求,大概需要一两天。 2025年6月10日龙虎榜行业方向分析 1. 金融科技
代表标的:御银股份、雄帝科技
驱动…
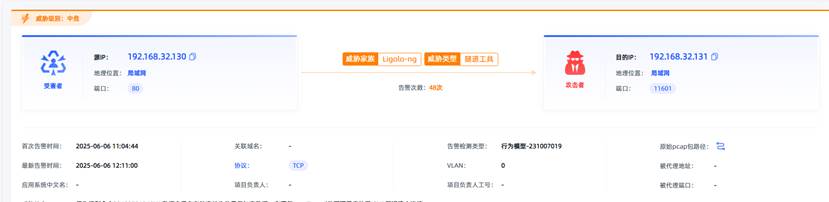
观成科技:隐蔽隧道工具Ligolo-ng加密流量分析
1.工具介绍
Ligolo-ng是一款由go编写的高效隧道工具,该工具基于TUN接口实现其功能,利用反向TCP/TLS连接建立一条隐蔽的通信信道,支持使用Let’s Encrypt自动生成证书。Ligolo-ng的通信隐蔽性体现在其支持多种连接方式,适应复杂网…

铭豹扩展坞 USB转网口 突然无法识别解决方法
当 USB 转网口扩展坞在一台笔记本上无法识别,但在其他电脑上正常工作时,问题通常出在笔记本自身或其与扩展坞的兼容性上。以下是系统化的定位思路和排查步骤,帮助你快速找到故障原因:
背景:
一个M-pard(铭豹)扩展坞的网卡突然无法识别了,扩展出来的三个USB接口正常。…

未来机器人的大脑:如何用神经网络模拟器实现更智能的决策?
编辑:陈萍萍的公主一点人工一点智能 未来机器人的大脑:如何用神经网络模拟器实现更智能的决策?RWM通过双自回归机制有效解决了复合误差、部分可观测性和随机动力学等关键挑战,在不依赖领域特定归纳偏见的条件下实现了卓越的预测准…

Linux应用开发之网络套接字编程(实例篇)
服务端与客户端单连接
服务端代码
#include <sys/socket.h>
#include <sys/types.h>
#include <netinet/in.h>
#include <stdio.h>
#include <stdlib.h>
#include <string.h>
#include <arpa/inet.h>
#include <pthread.h>
…

华为云AI开发平台ModelArts
华为云ModelArts:重塑AI开发流程的“智能引擎”与“创新加速器”!
在人工智能浪潮席卷全球的2025年,企业拥抱AI的意愿空前高涨,但技术门槛高、流程复杂、资源投入巨大的现实,却让许多创新构想止步于实验室。数据科学家…

深度学习在微纳光子学中的应用
深度学习在微纳光子学中的主要应用方向
深度学习与微纳光子学的结合主要集中在以下几个方向:
逆向设计 通过神经网络快速预测微纳结构的光学响应,替代传统耗时的数值模拟方法。例如设计超表面、光子晶体等结构。
特征提取与优化 从复杂的光学数据中自…