Nature Microbiology | 效应蛋白-宿主相互作用组图谱将健康肠道微生物组中的III型分泌系统与免疫调节联系起来
news2026/3/19 9:47:42
点击蓝字 关注我们Nature Microbiology | Pascal Falter-Braun 组-效应蛋白-宿主相互作用组图谱将健康肠道微生物组中的III型分泌系统与免疫调节联系起来研究论文DOIIF: 18.7 Q1 B1https://doi.org/10.1038/s41564-025-02241-y原文链接https://www.nature.com/articles/s41564-025-02241-y第一作者Veronika Young, Bushra Dohai通讯作者Andreas Zanzoni(andreas.zanzoniuniv-amu.fr), Pascal主要单位慕尼黑亥姆霍兹环境健康研究中心、艾克斯-马赛大学、乌得勒支大学医疗中心、丹娜-法伯癌症研究所、哈佛医学院、列日大学、慕尼黑大学、慕尼黑工业大学摘要Abstract假单胞菌门Pseudomonadota原变形菌门在人类肠道共生菌群中十分普遍但也包含许多病原体这些病原体依赖分泌系统将蛋白质注入宿主细胞以维持其致病性。在此我们表明来自健康肠道微生物组的假单胞菌门中有80%也具有完整的III型分泌系统T3SS。由机器学习预测的候选效应蛋白在序列和结构特征上表现出与病原体效应蛋白截然不同的特点。为了在系统层面上理解其功能我们通过实验构建了人类蛋白质与共生菌效应蛋白之间的蛋白质-蛋白质元相互作用组meta-interactome图谱。网络分析揭示效应蛋白靶向的邻近网络区域富含与微生物组相关疾病包括自身免疫性疾病和代谢性疾病相关的遗传变异。宏基因组学分析显示效应蛋白在克罗恩病中富集而在溃疡性结肠炎中则出现耗竭。在功能上共生菌效应蛋白能够转运到人类细胞中并在体外调节NF-κB信号传导和细胞因子的分泌。我们的研究结果表明T3SS有助于微生物与宿主的共生且效应蛋白-宿主蛋白质之间的相互作用可能是共生肠道菌群影响健康的一条被低估的途径。结果ResultsT3SS在人类肠道微生物组中很常见我们首先分析了假单胞菌门Pseudomonadota菌株的参考基因组这些菌株分离自健康肠道和粪便样本例如由人类微生物组计划分离。利用广泛用于分泌系统鉴定的工具EffectiveDB我们在77个基因组中的44个中检测到了完整的T3SS补充数据1。为了扩大研究范围我们分析了来自人类肠道细菌集合HiBC、Broad研究所-OpenBiome微生物组文库BIO-ML以及全球微生物组保护协会GMC的4,752个系统发育多样的菌株基因组。在568个假单胞菌门基因组中有449个79%具有完整的T3SS扩展数据图1类似比例的菌株具有T4SS315个和T6SS474个它们除了其他功能外也可能将效应蛋白注入宿主细胞扩展数据图1和补充数据2。总体而言在568个假单胞菌门基因组中有527个92%至少具有一种靶向宿主的分泌系统。由于培养可能会导致分类群代表性出现偏差我们还筛选了16,179个高质量和中等质量的假单胞菌门宏基因组组装基因组MAGs在770个5%MAGs中发现了完整的T3SS扩展数据图1和补充数据3。值得注意的是T3SS仅在γ-变形菌纲Gammaproteobacteria中被检测到而在β-或ε-变形菌纲中未检测到螺杆菌属菌株除外并且在埃希氏菌属Escherichia中尤为常见图1a和补充数据3。在T3SS阳性T3SS物种中有24个在魏茨曼科学研究所的两个队列中找到了匹配的代表物种以色列队列和荷兰队列中分别有59.4%和47.1%的个体在其肠道微生物组中携带潜在的T3SS物种其相对丰度分别为0.80%和0.48%其中大肠杆菌Escherichia coli最为常见。这些观察结果表明T3SS菌株是人类肠道菌群的常见成员这也促使我们进行了进一步的调查研究。图1 | 共生肠道细菌中的 T3SSa, 在人类肠道参考菌株和宏基因组组装基因组MAGs中编码完整T3SS的最丰富的属、种和基因组。b, 3,002个共生候选T3SS效应蛋白与来自致病菌的1,195个效应蛋白在不同比对覆盖率下的序列相似性左下。每个点代表一次成对的序列比对。点的颜色表示具有显著和不显著Jackhmmer结果的效应蛋白插图图例这表明了与病原体效应蛋白的同源性。边缘直方图显示了比对覆盖率顶部和序列相似性右侧的聚合分布颜色表示Jackhmmer结果。c, 左图FoldSeek分析中观察到的结构聚类数量红色箭头与该组同质或混合在灰色中的随机期望值的比较期望经验 P 0.0001n 10,000双侧标签置换检验。中图该组中一个聚类的示例结构小型网络是该组具有代表性的结构聚类中心为锚定结构相似结构通过连线连接。环形图在结构聚类组的所有同质或混合聚类中由颜色指示来源的蛋白质比例。d, 为后续功能分析选择的18种共生假单胞菌门Pseudomonadota菌株。数字表示在90%的序列长度上相互序列相似性 90% 的共有效应蛋白数量。e, 通过重组纳米荧光素酶的化学发光y轴检测野生型和ΔsctVT3SS缺陷型鼠伤寒沙门氏菌向HeLa细胞中注射的指定效应蛋白。对照病原体效应蛋白左来自鼠伤寒沙门氏菌的 sseJ (A0A0F6B1Q8)、sopA (Q8ZNR3) 和 pipB2 (A0A0F6B5HS)来自假结核耶尔森氏菌的 yopJ (A0A0N9NCU6)以及来自福氏志贺氏菌的 ipaH9.8 (Q8VSC3) 和 ospG (Q99PZ6)。SipA是作为参考的检测对照。星号表示野生型和ΔsctV阴性菌株之间具有统计学意义的显著差异双侧Wilcoxon检验5次生物学重复每次包含4次技术重复。f, 将肠道共生菌迟钝爱德华氏菌Edwardsiella tarda的效应蛋白注射到HeLa细胞中。在野生型和ΔsctV鼠伤寒沙门氏菌中测试的SipA分别用作阳性和阴性对照双侧Wilcoxon检验*P 0.05**P 0.001NS不显著7次生物学重复每次包含4次技术重复。所有图表的原始数据和精确的P值可见于补充信息中描述的补充数据1、3、5、6和10中。箱线图e,f显示了中位数中心线和四分位距IQR箱体须线延伸至 1.5×IQR 范围内的最小值和最大值。共生效应蛋白与已知病原体效应蛋白无关使用三种互补的机器学习模型在T3SS参考基因组中高置信度地预测出了3,002个候选效应蛋白以下简称菌株效应蛋白补充数据4并在770个T3SS MAGs中预测出了182个候选效应蛋白元效应蛋白补充数据4。由于T3SS效应蛋白传统上与致病性相关我们将这些候选效应蛋白与已知病原体的1,195个T3SS效应蛋白进行了比较。在3,002个菌株效应蛋白中仅有17个0.5%以及在182个元效应蛋白中仅有6个3%与病原体的效应蛋白表现出高度的序列相似性在≥90%的长度上具有≥90%的相似性补充数据5。为了寻找较弱的相似性我们使用jackhmmer针对来自UniRef90的约1.24亿个非冗余细菌序列进行了迭代搜索。然而即使采用这种高灵敏度的方法也仅有155个共生菌株效应蛋白约5%和42个元效应蛋白22.5%被发现与病原体效应蛋白具有显著的相似性图1b和扩展数据图1。由于即使序列存在差异效应蛋白在结构上也可能相关因此我们使用FoldSeek对AlphaFold预测的三级结构进行了聚类分析以进行结构比较。令人惊讶的是仅包含来自共生菌株或病原体的效应蛋白的同质聚类被高度过度代表而反映病原体和共生菌株效应蛋白共有结构的混合聚类II和III则被耗竭图1c和补充数据6P0.0001经验 P值。元效应蛋白完全与菌株效应蛋白聚类尽管这接近于随机预期。在改变FoldSeek参数以及仅考虑脊椎动物或人类病原体时所有结果均具有稳健性补充数据6。因此来自健康人类肠道T3SS菌株的候选效应蛋白在序列和结构上都与病原体效应蛋白存在显著差异。我们分析了来自菌株的所有候选效应蛋白的注释结构域。除了860个没有任何可识别结构域的蛋白质外最常见的发现包括双鸟苷酸环化酶GGDEF结构域PF0099058个效应蛋白和EAL结构域PF0056350个效应蛋白而这些在病原体效应蛋白中均未被发现补充数据5。环二鸟苷酸是细菌信号传导中一种已知的第二信使而EAL结构域被认为是双鸟苷酸磷酸二酯酶从而产生与环化酶相反的作用。此外我们在32个效应蛋白中观察到了PAS折叠结构域PF08447它可以作为配体结合传感器发挥作用并且在某些效应蛋白中与鸟苷酸环化酶结构域共存。由于环二核苷酸最近被认为是所有生命界中重要的免疫调节剂这两种作用于同一第二信使的结构域在共生候选效应蛋白中高频出现的观察结果使得这种信号分子在跨界通讯中发挥作用的可能性变得十分合理。将共生候选效应蛋白注入人类细胞一个关键问题是共生候选效应蛋白是否会通过T3SS注入人类细胞中。为了进行功能研究我们克隆了编码来自18个细菌菌株效应蛋白的开放阅读框ORFs图1d扩展数据图1补充表9以及补充数据7和8。生成的“人类微生物组效应蛋白ORFeome v1HuMEOme_v1”包含了910个经过序列验证的全长ORF代表了746个菌株效应蛋白和164个元效应蛋白补充数据7。克隆失败主要归因于PCR扩增失败且没有毒性迹象。以鼠伤寒沙门氏菌Salmonella enterica subsp. enterica sv. Typhimurium, S. Typhimurium为模型我们建立了一种基于纳米荧光素酶nano-luciferase的注射检测方法 将一个由11个氨基酸组成的Nano-Luc HiBiT标签融合到在细菌中表达的候选效应蛋白的C端。HeLa细胞稳定表达互补的LgBiT片段因此效应蛋白的注入能够重新组装成具有功能的纳米荧光素酶。在所有测试中通过引入T3SS缺陷型的ΔsctV突变体来确保检测的特异性。使用6个病原体效应蛋白进行的基准测试表明其中4个能够被有效转运。在来自11个菌株的97个受试候选效应蛋白中有32个被特异性且显著地注入图1e扩展数据图1补充表9和补充数据10。阳性对照的成功率略高这可能反映了系统发育的多样性以及缺失的分子伴侣chaperones和辅助因子cofactors。因此尽管不能排除一些错误的效应蛋白鉴定结果但总体而言我们的分析流程可靠地从健康人类肠道的共生菌株中鉴定出了真正的T3SS底物效应蛋白。接下来我们评估了共生菌中T3SS的功能性。在至少具有一个可通过T3SS注入的效应蛋白的11个菌株中有6个由于抗生素耐药性或转化失败而无法进行测试。虽然两株大肠杆菌E. coli未产生信号但巴斯德柠檬酸杆菌Citrobacter pasteurii和马赛植杆菌Phytobacter massiliensis偶尔出现信号表明T3SS存在零星激活。相比之下迟钝爱德华氏菌Edwardsiella tarda可重复且显著地将其四个受试效应蛋白中的三个注入了HeLa细胞图1f和补充数据10。值得注意的是其中只有一个Eta_3在沙门氏菌系统中也呈阳性这支持了这样一种观点即第一组实验中辅助因子的缺失可能导致了假阴性结果。总体而言这些数据表明来自健康人类肠道的菌株中存在功能性的T3SS并且能够将鉴定的效应蛋白递送至人类细胞中。微生物组-宿主蛋白质-蛋白质元相互作用组图谱接下来我们利用多重检测作图流程multi-assay mapping pipeline系统地绘制了共生效应蛋白与宿主蛋白质的物理相互作用网络以探索其潜在功能扩展数据图2。通过针对全人类ORFeome9.1筛选所有克隆的效应蛋白我们鉴定出了1,067个相互作用构成了人类-微生物组元相互作用组HuMMI的主数据集HuMMI_MAIN图2a。使用290个效应蛋白和1,440个人类蛋白质进行的独立重复筛选共三次产生了39个相互作用HuMMI_RPT并表明主筛选的采样灵敏度约为32%图2b这与先前的研究结果相一致。最后我们探讨了序列相似性如何影响效应蛋白的相互作用谱。我们将序列同一性≥30%的效应蛋白进行分组补充数据11并针对它们在主筛选中相互作用靶标的并集进行了实验验证测试。由此产生的HuMMI_HOM数据集包含394个相互作用其中181个为非冗余相互作用。总而言之HuMMI包含了286个效应蛋白与426个人类蛋白质之间的1,255个独特且经过验证的相互作用图2a和补充数据11。图2 | 细菌效应蛋白与人类蛋白质的元相互作用组网络图谱a, 经过验证的人类微生物组元相互作用组HuMMI图谱彩色节点表示来自不同菌株的效应蛋白颜色对应图f中的图例。灰色节点代表人类蛋白质外层的人类蛋白质仅被距离最近的菌株靶向中心的人类蛋白质则被来自多个菌株的效应蛋白靶向。b, 采样灵敏度根据 HuMMI_RPT 计算的饱和曲线。红点表示在指定数量的重复筛选的任意组合中发现的经过验证的相互作用的平均值菱形表示在所有连续实验组合中每次实验的相互作用计数误差线表示标准差standard deviation黑点和黑线表示计算得出的饱和曲线。。c, 检测灵敏度在用于网络作图的Y2H系统酵母双杂交系统 中来自 bhLit_BM-v1n 54对、bhRRS-v1n 72对、hsPRS-v2n 60对和 hsRRS-v2n 78对这几个集合中鉴定出的相互作用的百分比。误差线表示比例的标准误s.e. of proportion。d, 在 yN2H酵母纳米荧光素酶双杂交验证检测中HuMMI相互作用随机样本n 294种配对组合与四个参考集的验证率比较bhLit_BM-v1n 94种配对组合bhRRS-v1n 144种配对组合hsPRS-v2n 44种配对组合和 hrRRS-v2n 51种配对组合。双侧 Fisher 精确检验*P 0.04***P 0.0006补充数据14。误差线表示比例的标准误。e, 带 Flag 标签的效应蛋白或作为阴性对照的 Flag-GFP 对带 MYC 标签的人类蛋白质的免疫共沉淀Co-immunoprecipitation。Input 代表细胞裂解液绿点表示成功的免疫共沉淀红点表示无免疫共沉淀大肠杆菌Escherichia coli的效应蛋白 espGQ7DB50作为阳性对照一次生物学重复。分子量标记Molecular mass markers的单位为千道尔顿kDa。f, 与指定数量的来自不同菌株的效应蛋白发生相互作用的最常被靶向的人类蛋白质。颜色代表不同菌株对应指定的图例完整统计信息见补充数据11。g, 与指定数量的人类蛋白质发生相互作用的连接度connectivity最高的效应蛋白补充数据11。h, 观察到的与效应蛋白发生相互作用的人类蛋白质数量与随机预期值的比较双侧置换检验P 0.0001n 10,000。i, 被指定数量的不同菌株的效应蛋白靶向的人类蛋白质的频率分布红色与随机预期值的比较双侧置换检验P 0.004n 10,000。为了通过实验评估数据质量我们组装了一个由67个记录良好的病原体效应蛋白与人类蛋白质二元相互作用组成的阳性对照集细菌人类文献二元多重集bhLit_BM-v1以及一个由随机效应蛋白-人类蛋白质对组成的阴性对照集细菌宿主随机参考集bhRRS-v1补充数据12。使用这些对照集以及已建立的人类参考集hsPRS-v2和hsRRS-v2对我们的酵母双杂交Y2H检测 进行基准测试结果显示检测灵敏度分别为13%和17.5%这与先前的观察结果相一致图2c和补充数据13。没有任何阴性对照得分为阳性这证明了我们系统的可靠性。接下来我们使用参照这四个参考集进行基准测试的酵母纳米荧光素酶双杂交检测yN2H评估了HuMMI的生物物理质量。在不同的阈值下包含细菌蛋白质的集合产生的阳性得分对少于人类集合扩展数据图2。由于这包含了阴性对照并且没有观察到效应蛋白具有毒性原核蛋白质似乎在这种检测系统中更难进行测试这进一步强化了对定制参考集的需求。随机选择的172个HuMMI相互作用在统计学上与阳性对照集无法区分图2d扩展数据图2和补充数据14。因此HuMMI的生物物理质量与文献中充分记录的相互作用相当。我们的目标是证明这些相互作用可以发生在人类细胞环境中。我们在HEK293细胞RRIDCVCL_0045DSMZ中进行了免疫沉淀实验 使用带Flag标签的效应蛋白和作为阴性对照的Flag-GFP作为诱饵baits并通过蛋白质印迹法Western blot检测带有MYC标签的人类相互作用伙伴。在包含4个阳性对照的32个测试对中18对候选对和3个对照产生了有意义的数据而有10对由于人类蛋白质的非特异性结合3对或表达不良7对而无法评估。对照对中只有一对呈阳性而18个候选对中有13个在效应蛋白免疫沉淀中特异性地产生了可检测的条带图2e和扩展数据图2。综合来看这些结果表明HuMMI包含了生物物理学上可靠的相互作用这些相互作用在不同的检测方法中都能被稳健地检测到并且能够真实发生于人类细胞环境中。重要的是功能性影响可能是双向的虽然在大多数情况下效应蛋白可能会扰乱宿主细胞但细胞内免疫受体也可能识别效应蛋白并随后启动防御反应。我们通过分析微生物-宿主相互作用网络的拓扑结构开始了功能探索图2fg。HuMMI_MAIN的度分布degree distribution显示大量人类蛋白质与多个效应蛋白通常来自不同物种存在相互作用图2f和补充数据11。随机抽样分析表明效应蛋白在少数宿主相互作用蛋白上表现出极其显著的趋同性convergence图2h正如在植物-病原体系统中所展示的那样这种现象与被靶向的宿主蛋白在功能上的重要性有关。此外人类蛋白质与来自四个细菌菌株的效应蛋白的相互作用非常显著不太可能是由随机过程引起的图2i和补充数据11。因此有60个人类蛋白质受到效应蛋白趋同靶向的影响突显了它们在微生物组-宿主相互作用中的潜在重要性。为了探索与病原体效应蛋白的重叠部分我们从IntAct数据库中提取了来自17个致病菌株的80个效应蛋白与217个人类蛋白质之间的265个高质量二元相互作用补充数据15。我们发现两组共生菌与病原体共同靶向的人类蛋白质数量虽然在数值上较低但具有显著性12个人类蛋白质P 0.014Fisher精确检验比值比 2.26其中有3个蛋白质受到了共生效应蛋白的趋同靶向P 0.067Fisher精确检验比值比 3.37补充数据11。尽管受到样本量、实验差异和病原体数据非系统性特征的限制这些发现依然支持了共生菌和病原体在效应蛋白靶向机制中既存在重叠又具有依赖于生活方式致病或共生的特异性。介导效应蛋白-宿主相互作用的结构特征许多推断方法假设序列相似性意味着功能和相互作用的相似性并且这种相似性也可能是趋同convergence的基础。然而在经过系统测试的 HuMMI_HOM 数据集的同源聚类homology clusters中图3a 我们发现序列相似性与相互作用相似性之间的相关性很差相反序列相似性仅仅界定了相互作用相似性的上限。例如聚类3包含7个序列相似性大于90%的效应蛋白但它们的相互作用谱相似性却表现出从完全相同到互补complementary的广泛分布图3b,c和补充数据16。反之在 HuMMI_MAIN 中根据成对相互作用的相似性对那些在序列和结构上毫不相关的效应蛋白进行聚类发现在同源聚类之外存在实质性的相互作用靶标重叠扩展数据图3这表明不相似的效应蛋白在宿主体内可能具有相似的功能。因此以蛋白质相互作用谱为衡量标准的宿主效应蛋白功能在很大程度上独立于整体序列的相似性。图3 | 相互作用特异性与相互作用基序a, 同源效应蛋白系统性相互作用谱分析的模式图。b, 指定同源组中所有效应蛋白对的相互序列相似性与 Jaccard 相互作用相似性的散点图。每对效应蛋白靶向的人类蛋白质并集由节点大小表示如图例所示。个体数据见补充数据16。c, 针对同源聚类3homology cluster 3测试的所有效应蛋白-人类蛋白质相互作用的四次重复实验中其中一次代表性实验的酵母生长情况。d, 左图与同一细菌效应蛋白发生相互作用的人类蛋白质靶标的比例根据靶标接触残基的 Jaccard 指数JI计算的界面相似性进行分组分类为截然不同distinctJI ≤ 0.1、重叠overlapping0.1 JI 0.6和相同sameJI ≥ 0.6。右图Mmo_5 通过相同的界面顶部66% 重叠与示例人类蛋白质相互作用而 Pfa_4 则使用截然不同的界面。e, 与同一人类蛋白质发生相互作用的不同效应蛋白的比例按与图d相同的界面相似性标准进行分组。示例界面模型右显示效应蛋白结合在人类 TCF4 上相同Pfa_9Pse_2或截然不同Pfa_9Yen_2的界面上。f, 在 HuMMI_MAIN 中鉴定出的满足至少一项严谨标准箭头所示的结构域-基序domain–motif界面计数与随机预期值的比较单侧置换检验P 0.0137n 10,000。与c-f相关的全部数据可见于补充数据17。g, Holdup 检测holdup assay结果及其与酵母双杂交Y2H结果的比较。 针对顶部标示的效应蛋白的10个氨基酸组成的C端多肽对y轴上显示的人类蛋白质指定 PDZ 结构域进行了结合测试。计算得出的解离常数Kd值如标示所示。彩色边框表示 HoldupHU和 Y2H 在蛋白质水平上的相互作用重叠。每次测试的精确P值和n值显示在补充数据19中。为了获得结构上的见解和潜在的功能线索我们使用 AlphaFold-Multimer 对效应蛋白-宿主蛋白质的相互作用进行了建模 获得了 123 对10%的预测结果。对于具有多个相互作用伙伴的蛋白质我们将结合界面分类为“相同”≥60% 的共享接触残基、“不同”10% 的重叠或“重叠”图3d,e 和扩展数据图3。例如Mmo_5 通过相同的界面与 TPD52L1 和 BORCS6 结合而 Pfa_4 则通过不同的界面与 NOTO 和 LBX1 相互作用这使得其可能与两者同时发生相互作用图3d 和补充数据17。类似地Pse_2 和 Pfa_9 结合 TCF4 的相同界面而 Yen_2 则靶向该蛋白质的不同部分图3e。相同界面的结合在人类蛋白质上比在效应蛋白上更常见这表明靶向与特定结构域相关的功能具有重要意义。将结合界面映射到结构域注释强化了这一假设因为即使是通过不同界面结合的效应蛋白也可能靶向相同的结构域例如LBX1 的 DNA 结合结构域。然而更常见的情况是具有不同界面的效应蛋白会结合到宿主蛋白质的不同部位。Efe_11 和 Kpn_9 结合在 TRAF2 的 E3 泛素连接酶结构域的相同界面上而 Pem_8 则靶向 TRAF2 的 C 端 MATH 结构域该结构域介导三聚化和受体结合。同样在 REL 蛋白上Pma_4 结合 DNA 结合结构域而 Yen_11 则结合二聚化结构域。除了大型界面外许多相互作用是由本质无序区域中的短线性基序SLiMs介导的这些基序结合到特定的形成口袋的蛋白质结构域上。由于 AlphaFold 经常遗漏此类相互作用我们使用了正交的 mimicINT 方法来鉴定 SLiM-结构域的相互作用该方法将相互作用对与已知的 SLiM-结构域模板进行匹配图3f。这为涉及细菌类宿主 SLiMs 结合到人类结构域的 54 个 HuMMI_MAIN 相互作用确定了推定的界面补充数据18其中 51 个满足了至少一项严格标准图3fP 0.0137经验 P 值22个满足了两项严格标准扩展数据图3P 0.0005经验 P 值。一些匹配的基序包含与激酶或依赖磷酸化的结合结构域如 SH2 结构域发生相互作用的磷酸化位点。相反尽管几个共生效应蛋白编码了预测的酶结构域补充数据5但使用类似的方法我们没有发现这些结构域结合宿主蛋白质上同源底物基序的案例并且只有一个符合已知对接特异性的效应蛋白结构域-SLiM 匹配Efe_1 的类钙调磷酸酶磷酸酯酶结构域PF00149和 VAC14 中的经典 LxVP 对接基序。最大的一组共 23 个相互作用涉及人类蛋白质中的 PDZ 结构域与细菌相互作用伙伴 C 端的 PDZ 结合基序PBM的结合。含有 PDZ 结构域的蛋白质通常介导对微生物-宿主相互作用非常重要的功能包括细胞-细胞黏附、蛋白质运输和免疫信号传导。为了通过实验验证这些界面我们通过体外定量相互作用 holdup 检测方法holdup assay对来自 13 个人类蛋白质的单个和串联 PDZ 结构域以及来自 16 个相互作用细菌效应蛋白的 C 端多肽进行了测试。在 23 个酵母双杂交Y2H对中16 个70%显示出至少一种 PDZ-多肽相互作用从而验证了这种相互作用模式图3g 和补充数据19。在三个案例中发生相互作用需要两个串联排列的 PDZ 结构域这表明某些 Y2H 相互作用对可能由于未测试的组合而被 holdup 方法遗漏。至于预测的球状界面对于具有多个 PDZ 结构域的人类蛋白质不同的效应蛋白通常靶向不同的结构域这证明了其特异性和功能特化图3g。因此虽然效应蛋白的整体序列相似性与相互作用谱不相关但结构建模表明一些效应蛋白靶向相似的界面和结构域暗示具有共有功能而另一些效应蛋白则结合截然不同的结构域这指向了功能特化functional specialization。效应蛋白靶向的功能与疾病模块我们利用基因本体论GO富集分析探索了效应蛋白的靶标功能图4a扩展数据图3和补充数据20。在最显著富集的功能中包括“对胞壁酰二肽MDP的反应”MDP是一种源自细菌细胞壁的肽。有趣的是MDP的受体NOD2是克罗恩病Crohns disease的一个主要易感基因克罗恩病是一种肠道自身免疫性疾病微生物组在其病因学中起着重要作用。中枢免疫信号通路也显著富集即NF-κB通路以及应激激活蛋白激酶和c-Jun氨基末端激酶SAPK/JNK通路。值得注意的是有五个被显著靶向的趋同convergence蛋白质属于NF-κB模块扩展数据图3这是动物进化上最古老的免疫通路之一。使用 Recon3D 人类基因组规模代谢模型我们进一步发现在人类相互作用蛋白中代谢酶显著富集P 0.0001Fisher精确检验然而除了甘油磷脂代谢外没有其他代谢子系统表现突出补充数据20。最后我们将共生菌靶向的功能与病原体靶向的功能进行了比较补充数据20。某些通路是两组靶标所共有的例如“NF-κB信号传导”而另一些通路则特异性地属于共生菌包括“胶原生物合成”和“对胞壁酰二肽的反应”。这些发现强化了这样一种观念共生菌和病原体在与人类宿主的分子相互作用中既存在依赖于其生活方式致病或共生的特异性又存在功能上的重叠。图4 | 微生物靶向的人类蛋白质的功能与疾病关联a 在效应蛋白靶向的人类蛋白质中富集的代表性功能注释的比值比odds ratiosFDR 0.05带有 Bonferroni FDR 校正的 Fisher 精确检验。术语旁边的数字#显示了该类别所包含的术语数量。柱状图的浅色阴影区域表示在该代表组中观察到的最低和最高比值比。黑线表示所示代表性术语的比值比。白色三角形表示在病原体靶标中同样富集的功能。b 在人类参考相互作用组HuRI中编码效应蛋白相互作用蛋白的人类基因中显著富集的性状和疾病的遗传易感性genetic predispositionα 0.05Fisher 精确检验n 349。a 和 b 中的比值比评估了显著功能/性状的效应大小双侧 Fisher 精确检验 FDR 0.05其计算方法是在靶标集中编码效应蛋白靶标的带有功能注释/性状相关的人类基因与无功能注释/性状无关的人类基因的比率odds除以 HuRI 集中的相同比率详见“方法”部分。c 遗传易感性蛋白质在指定菌株效应蛋白的网络邻域network neighbourhoods中显著富集的疾病组。性状节点的大小对应于图例中指示的该组内被显著靶向的性状数量。菌株-分组之间连线的粗细反映了潜在的显著效应蛋白-性状关联的数量α 0.01 且比值比 3双侧 Fisher 精确检验。d c图“免疫immunological”组中所包含的具体疾病。节点大小反映了图例中指示的潜在效应蛋白-性状关联的数量。所有测试的精确 $P$ 值和样本量$n$均提供在补充数据23中。我们想知道共生效应蛋白的扰动是否会影响非传染性人类疾病并从网络层面开始了我们的分析。遗传变异以及病毒通常通过微妙地改变细胞内网络和与疾病相关的功能来促发复杂疾病。我们首先探讨了共生效应蛋白是否靶向在遗传学上与疾病和其他性状相关的蛋白质。我们使用了由 Open Targets 计划从全基因组关联分析GWAS 中鉴定出的“致病基因causal genes”来识别其编码的“疾病蛋白质disease proteins”并通过实验因子本体论EFO统一了这些性状图4b 和补充数据21。“免疫球蛋白同种型转换immunoglobulin isotype switching”的强烈富集现象非常有趣因为在进化上较古老的 IgA 抗体在塑造肠道微生物组方面发挥着重要作用。效应蛋白靶标也与癌症和免疫疾病如银屑病、哮喘、过敏和系统性红斑狼疮相关尽管这些疾病主要影响的都不是肠道。鉴于存在大量免疫相关的测量性状效应蛋白有可能系统性地扰动免疫信号传导从而促发肺部和皮肤疾病。或者诸如 REL 或 TCF4图2f,h等趋同convergence蛋白质也可能被皮肤或肺部组织中的局部微生物群所靶向。支持这一观点的是在皮肤微生物组样本中可检测到 26% 的 HuMMI 效应蛋白这表明共生效应蛋白在不同的生态位中是共享的补充数据22。除了疾病蛋白质作为直接靶标外我们之前还发现相关的遗传变异通常存在于它们的蛋白质相互作用邻域protein interaction neighbourhood中。为了探索这些邻域我们在二元人类参考相互作用组HuRI中执行了短随机游走short random walks并将“邻域”定义为在 HuRI 中被访问的频率显著高于在保持度数degree-preserved不变的随机重连对照网络中被访问频率的蛋白质。在这些邻域中我们使用 Open Targets 评估了疾病蛋白质的富集情况在菌株水平上汇总了名义显著的nominally significant关联并按疾病组对它们进行了总结图4c 和补充数据23。我们发现的大多数疾病组已知都受肠道微生物组的影响。在免疫疾病中炎症性肠病IBD显著富集名义 P 0.0008Fisher 精确检验特别是克罗恩病名义 P 8.5 × 10⁻⁵Fisher 精确检验但溃疡性结肠炎则未见富集图4d 和补充数据23。与直接靶标类似这些邻域也包含了对皮肤和肺部疾病如哮喘和银屑病的易感性。考虑到微生物群与代谢紊乱的相关性效应蛋白靶向影响高密度和低密度脂蛋白分别为 HDL 和 LDL胆固醇水平的邻域分别为名义 P 0.006 和 P 0.008Fisher 精确检验以及几个与糖尿病相关的性状这一点非常值得注意补充数据23。综合来看这些发现表明共生效应蛋白可调节宿主的免疫信号传导以及局部的代谢和结构微环境。由于靶向的蛋白质及其邻域在遗传上与多种疾病相关效应蛋白对其功能的调节可能会促成疾病的病因学disease aetiology。效应蛋白在人类细胞及疾病中的功能我们试图通过实验验证共生效应蛋白会扰动某些已鉴定的通路和功能。我们将重点放在了 NF-κB 信号传导上它是许多疾病的核心并在我们的研究中反复出现。使用 HEK293 细胞RRIDCVCL_0045DSMZ中的双荧光素酶报告基因检测 在无刺激的条件下26个共生效应蛋白中有5个显著激活了 NF-κB 活性图5a,b 和补充数据24而3个效应蛋白在强烈的 TNF 刺激下降低了 NF-κB 活性图5b扩展数据图4 和补充数据24。接下来我们评估了这些效应蛋白是否在未受刺激的 Caco-2 细胞中以及在促炎刺激后调节 NF-κB 信号传导扩展数据图4。一致的是包含 ABC 结构域的效应蛋白 Cpa_12在有刺激和无刺激的情况下均减少了几种细胞因子的分泌图5c 和补充数据25。其他效应蛋白仅在 Pam3CSK4 刺激后增强了细胞因子反应特别是 IL-6 和 IL-8但在 TNF 或鞭毛蛋白flagellin刺激后则没有增强图5d扩展数据图4 和补充数据25。Pam3CSK4 模拟了富含于革兰氏阳性拟杆菌门Bacteroidetes中的三酰化脂肽对 TLR1/2 的激活而鞭毛蛋白模拟了革兰氏阴性假单胞菌门Pseudomonadota对 TLR5 的激活。因此共生效应蛋白对细胞内免疫信号传导发挥着复杂的影响。图5 | 效应蛋白对人类细胞功能的影响及其在炎症性肠病IBDs中的临床流行率(a 表达指定效应蛋白的 HEK293 细胞在基线条件无 TNF 刺激下的相对 NF-κB 转录报告基因活性采用 Kruskal-Wallis 检验及 Dunn 事后多重比较*P 0.05**P 0.01n 4 个生物学重复。箱体表示四分位距IQR黑线表示平均值须线表示在 1.5× IQR 范围内的最高和最低数据点。b 效应蛋白在基线条件-TNF和 TNF 刺激后TNF对归一化 NF-κB 转录报告基因活性的显著影响总结采用 Kruskal-Wallis 检验及 Dunn 事后多重比较*P 0.05**P 0.01n 4 个生物学重复。c, d 转染了指定效应蛋白的 Caco-2 细胞在基础条件未刺激unstim或受到促炎混合物刺激stimc或受到 Pam3CSK4 刺激d后分泌的细胞因子浓度。EV 代表空载体模拟对照empty vector mock control。括号上方的数字表示通过 Kruskal-Wallis 检验及 Dunn 事后多重比较计算的 P 值n 3c和 n 5d个生物学重复。箱体表示 IQR黑线表示平均值圆圈表示个体数据点。e 放射状柱形图Radial barplot显示了 122 个细菌效应蛋白在克罗恩病CDn 504 个患者样本橙色或溃疡性结肠炎UCn 302 个患者样本紫色患者宏基因组中相对于健康对照组n 334 个样本的流行率prevalence倍数变化fold change。倍数变化是使用来自健康对照组的伪计数pseudo-counts计算的补充数据 26。标签指示了在克罗恩病或溃疡性结肠炎中具有显著流行率的效应蛋白FDR 0.01带有 Benjamini-Hochberg (BH) 校正的 Fisher 精确检验。黑色星号标记了各个柱子的统计学显著性。f 与健康对照组相比指定效应蛋白在克罗恩病n 504和溃疡性结肠炎n 302患者宏基因组中的流行率FDR 0.01双侧 Fisher 精确检验BH 校正。g HuMMI 子网络subnetworks显示了与克罗恩病橙色边框或溃疡性结肠炎紫色边框相关的人类蛋白质灰色节点它们与来自患者宏基因组中富集或耗竭菌株的效应蛋白彩色节点发生相互作用。效应蛋白颜色如图例所示。连线Edges代表 HuMMI 中的蛋白质-蛋白质相互作用。鉴于共生效应蛋白与 IBD 之间存在遗传和功能上的联系我们想知道临床数据是否支持效应蛋白在该疾病中的潜在作用。假设效应蛋白在 IBD 病因学中的因果关联可能反映在效应蛋白流行率prevalence的改变上我们分析了一项包含 800 多名 IBD 个体504 名克罗恩病患者302 名溃疡性结肠炎患者和 334 名健康对照者的宏基因组研究。聚焦于在 HuMMI 中具有物理相互作用的效应蛋白与健康对照组相比64个效应蛋白在克罗恩病患者中的流行率显著更高而这些效应蛋白在溃疡性结肠炎患者中则较不常见图5e,f 和补充数据26。这种相反的趋势出乎意料因为据报道假单胞菌门Pseudomonadota的整体丰度abundance在两种 IBD 中均有所增加。从 HuMMI 网络中得出了一些解释这一观察结果潜在机制的假设在克罗恩病中高度流行的肺炎克雷伯菌K. pneumoniae、大肠杆菌E. coli和弗格森大肠杆菌E. fergusonii的效应蛋白与克罗恩病易感蛋白 COG6 发生相互作用而 COG6 直接与溃疡性结肠炎易感基因 RTP5 发生相互作用图5g。类似地弗格森大肠杆菌的 Efe_13 结合克罗恩病易感蛋白 TNIP1该蛋白在 NF-κB 信号传导中发挥作用并与两个与溃疡性结肠炎相关的基因发生相互作用。其他富集的效应蛋白通过共有的相互作用伙伴显示出与克罗恩病和溃疡性结肠炎相关蛋白质的间接联系图5g。虽然这些相互作用的具体机制相关性还需要未来的实验研究来证实但这些与 IBD 疾病蛋白质的直接和间接分子网络联系引发了推测即它们可能导致了宿主内环境稳态的偏移homeostatic shift从而增加了克罗恩病的发病风险同时降低了溃疡性结肠炎的风险。作者简介Veronika Young与 Bushra Dohai 第一作者第一作者德国慕尼黑亥姆霍兹中心网络生物学研究所 Veronika Young 博士与 Bushra Dohai 博士为该论文的共同第一作者 慕尼黑亥姆霍兹中心网络生物学研究所INET所长、路德维希-马克西米利安-慕尼黑大学LMUPascal Falter-Braun 教授与艾克斯-马赛大学 Andreas Zanzoni 博士为论文的共同通讯作者。Andrew J. Macpherson 通讯作者Pascal Falter-Braun 是德国慕尼黑亥姆霍兹中心Helmholtz Munich网络生物学研究所INET的所长并担任路德维希-马克西米利安-慕尼黑大学LMU微生物-宿主相互作用主席。其实验室致力于利用系统生物学、高通量蛋白质组学和计算建模等前沿工具研究跨界分子相互作用网络特别是人类宿主与病原体及共生微生物之间的蛋白质-蛋白质“元相互作用组”meta-interactome。其研究核心在于揭示微生物如何利用分泌系统如 III 型分泌系统 T3SS将效应蛋白注入宿主细胞进而精确调节宿主的免疫信号传导如 NF-κB 通路。通过整合比较基因组学与结构预测AlphaFold其实验室不仅发现了共生菌中普遍存在的功能性分泌系统还揭示了这些相互作用如何影响自身免疫性疾病如克罗恩病和代谢疾病的易感性。Pascal 教授早年曾在哈佛医学院和丹娜-法伯癌症研究所工作 在构建人类参考相互作用组图谱方面做出了重要贡献。其学术成果广泛发表于 Nature、Science、Cell 和 Nature Microbiology 等国际顶级期刊。来源https://www.helmholtz-munich.de/en/inet/pascal-falter-braun翻译于乔西北农林科技大学硕士在读审核朱志豪广东医科大学基因组所联合博士后终审刘永鑫中国农科院基因组所研究员/博导排版申子昂华南理工大学基因组所联培硕士在读宏基因组推荐4月10-12日微生物组-扩增子16S分析5月8-10日微生物组-宏基因组分析本公众号现全面开放投稿希望文章作者讲出自己的科研故事分享论文的精华与亮点。投稿请联系小编微信号yongxinliu 或 meta-genomicsiMeta高引 fastp PhyloSuite ImageGP2 iNAP2 ggClusterNet2iMeta工具 SangerBox2 美吉2026 OmicStudio Wekemo OmicShareiMeta综述 高脂饮食菌群 发酵中药 口腔菌群 微塑料 癌症 宿主代谢10000扩增子EasyAmplicon 比较基因组JCVI 序列分析SeqKit2 维恩图EVenniMetaOmics高引 猪微生物组 16S扩增子综述 易扩增子(EasyAmplicon)系列教程微生物组入门 Biostar 微生物组 宏基因组专业技能学术图表 高分文章 生信宝典 不可或缺的人点击阅读原文
本文来自互联网用户投稿,该文观点仅代表作者本人,不代表本站立场。本站仅提供信息存储空间服务,不拥有所有权,不承担相关法律责任。如若转载,请注明出处:http://www.coloradmin.cn/o/2425970.html
如若内容造成侵权/违法违规/事实不符,请联系多彩编程网进行投诉反馈,一经查实,立即删除!相关文章
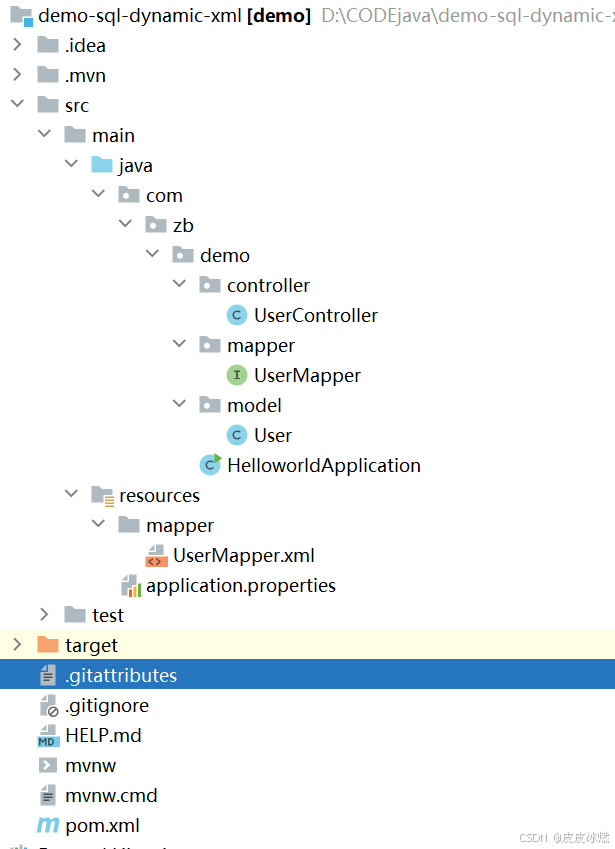
SpringBoot-17-MyBatis动态SQL标签之常用标签
文章目录 1 代码1.1 实体User.java1.2 接口UserMapper.java1.3 映射UserMapper.xml1.3.1 标签if1.3.2 标签if和where1.3.3 标签choose和when和otherwise1.4 UserController.java2 常用动态SQL标签2.1 标签set2.1.1 UserMapper.java2.1.2 UserMapper.xml2.1.3 UserController.ja…
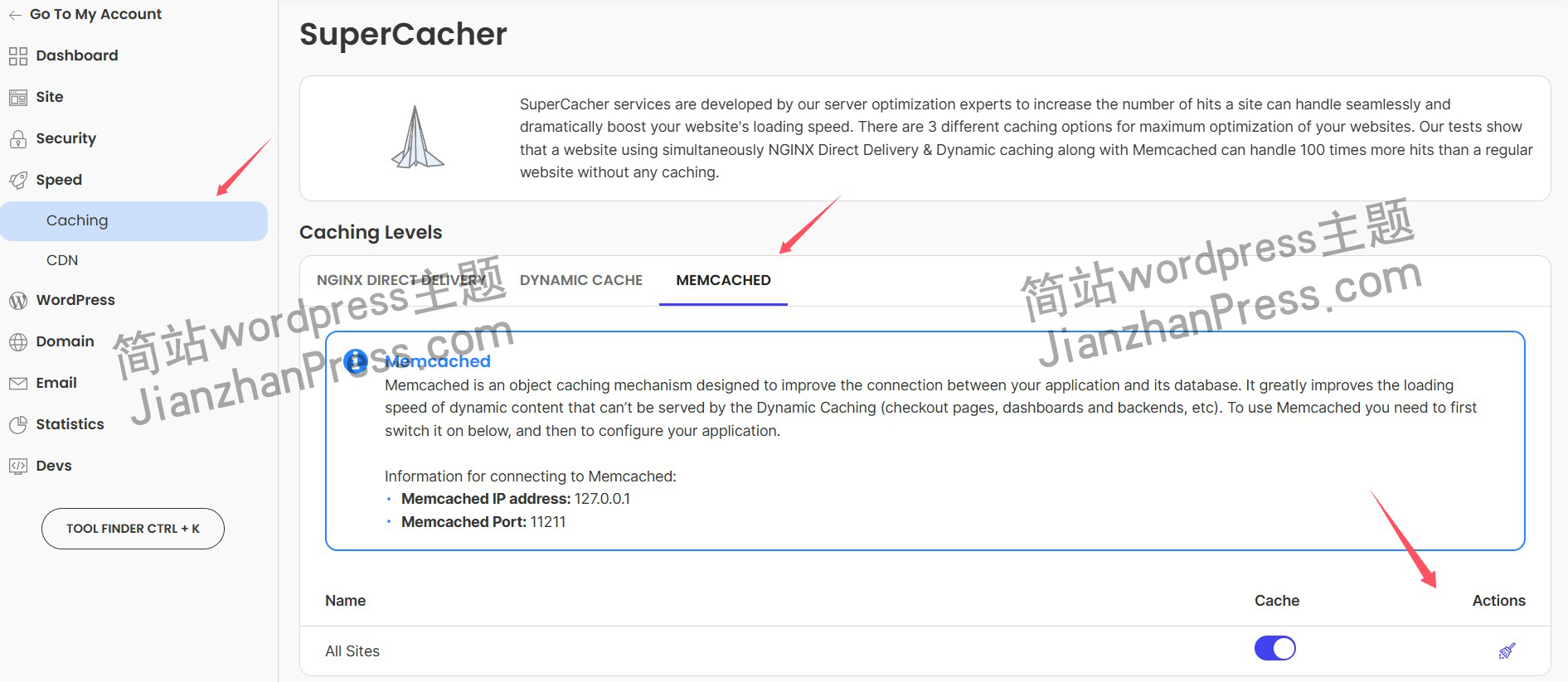
wordpress后台更新后 前端没变化的解决方法
使用siteground主机的wordpress网站,会出现更新了网站内容和修改了php模板文件、js文件、css文件、图片文件后,网站没有变化的情况。
不熟悉siteground主机的新手,遇到这个问题,就很抓狂,明明是哪都没操作错误&#x…
网络编程(Modbus进阶)
思维导图 Modbus RTU(先学一点理论)
概念 Modbus RTU 是工业自动化领域 最广泛应用的串行通信协议,由 Modicon 公司(现施耐德电气)于 1979 年推出。它以 高效率、强健性、易实现的特点成为工业控制系统的通信标准。 包…
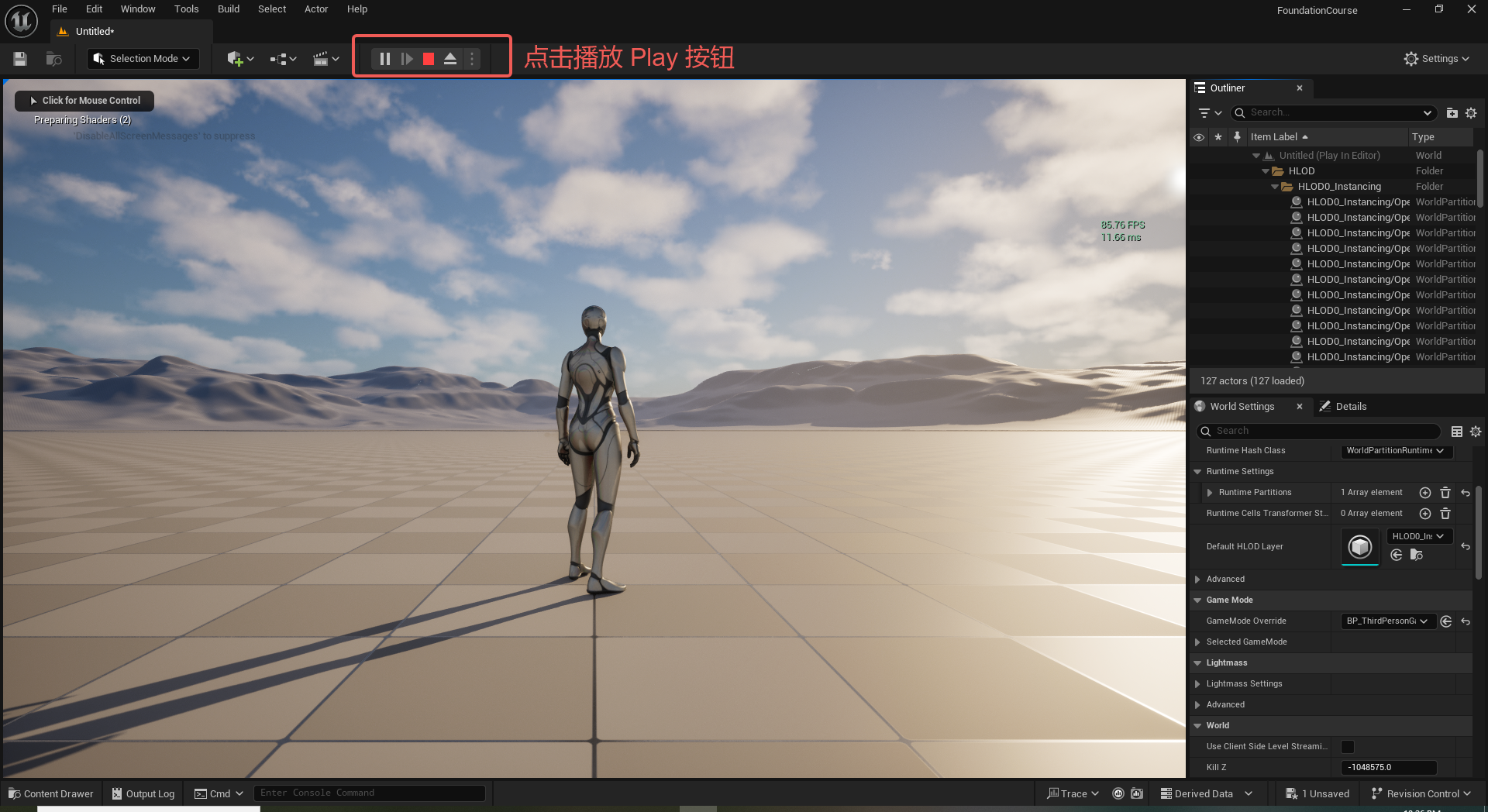
UE5 学习系列(二)用户操作界面及介绍
这篇博客是 UE5 学习系列博客的第二篇,在第一篇的基础上展开这篇内容。博客参考的 B 站视频资料和第一篇的链接如下:
【Note】:如果你已经完成安装等操作,可以只执行第一篇博客中 2. 新建一个空白游戏项目 章节操作,重…
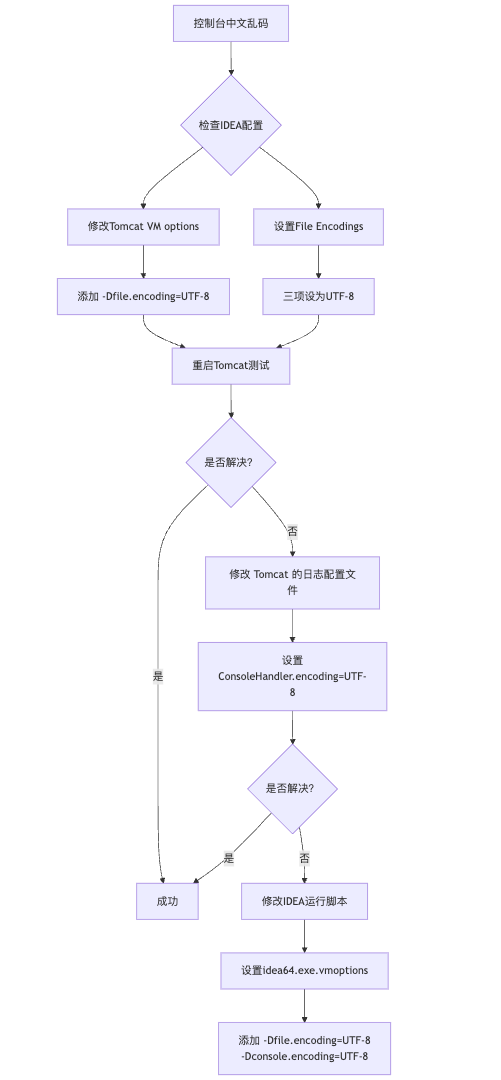
IDEA运行Tomcat出现乱码问题解决汇总
最近正值期末周,有很多同学在写期末Java web作业时,运行tomcat出现乱码问题,经过多次解决与研究,我做了如下整理:
原因:
IDEA本身编码与tomcat的编码与Windows编码不同导致,Windows 系统控制台…

利用最小二乘法找圆心和半径
#include <iostream>
#include <vector>
#include <cmath>
#include <Eigen/Dense> // 需安装Eigen库用于矩阵运算 // 定义点结构
struct Point { double x, y; Point(double x_, double y_) : x(x_), y(y_) {}
}; // 最小二乘法求圆心和半径 …
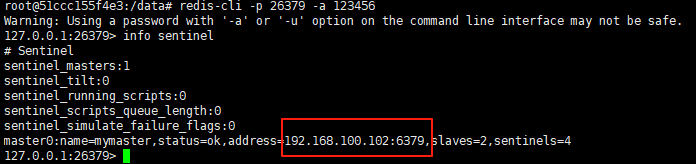
使用docker在3台服务器上搭建基于redis 6.x的一主两从三台均是哨兵模式
一、环境及版本说明
如果服务器已经安装了docker,则忽略此步骤,如果没有安装,则可以按照一下方式安装: 1. 在线安装(有互联网环境): 请看我这篇文章 传送阵>> 点我查看 2. 离线安装(内网环境):请看我这篇文章 传送阵>> 点我查看
说明:假设每台服务器已…
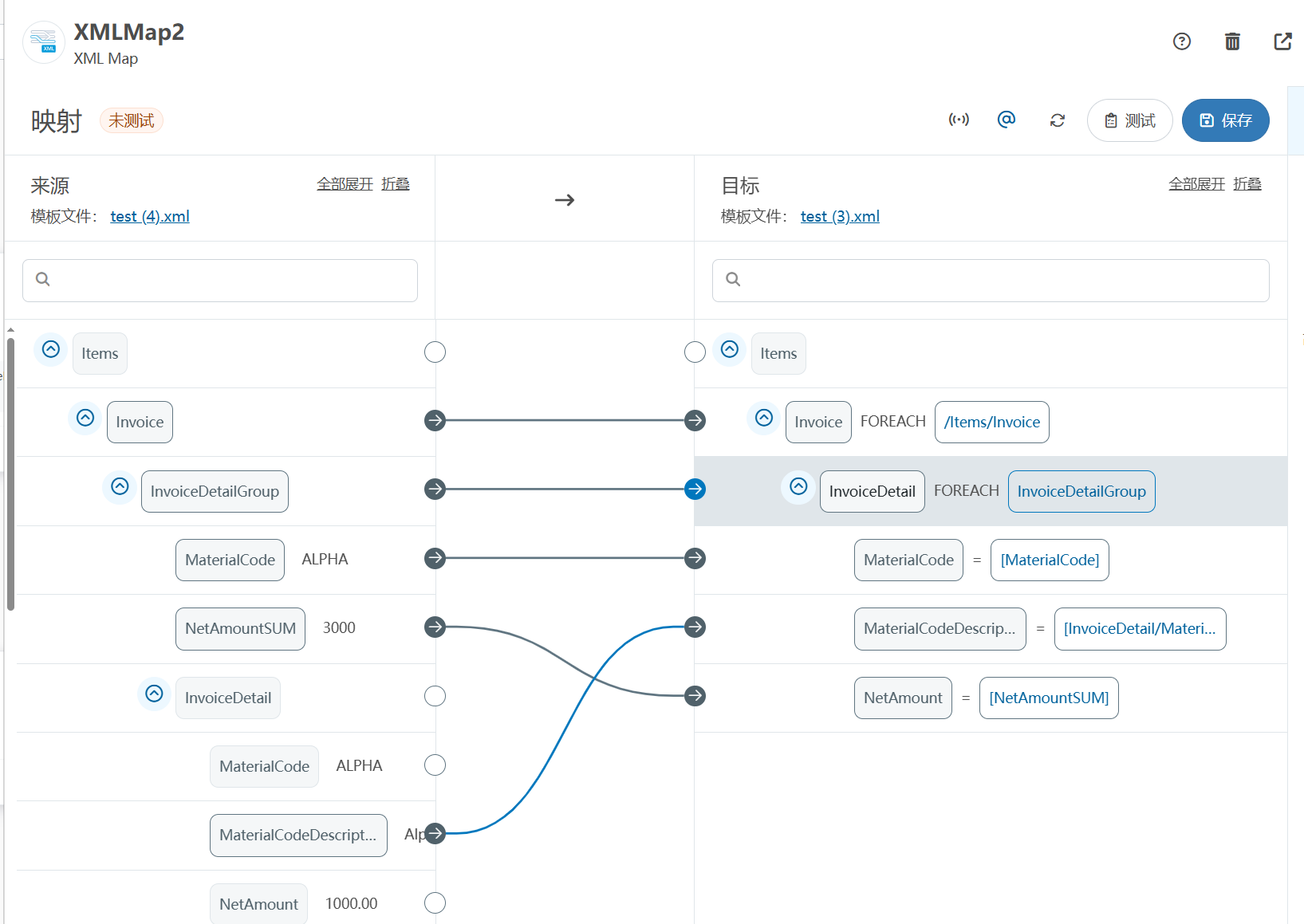
XML Group端口详解
在XML数据映射过程中,经常需要对数据进行分组聚合操作。例如,当处理包含多个物料明细的XML文件时,可能需要将相同物料号的明细归为一组,或对相同物料号的数量进行求和计算。传统实现方式通常需要编写脚本代码,增加了开…
LBE-LEX系列工业语音播放器|预警播报器|喇叭蜂鸣器的上位机配置操作说明
LBE-LEX系列工业语音播放器|预警播报器|喇叭蜂鸣器专为工业环境精心打造,完美适配AGV和无人叉车。同时,集成以太网与语音合成技术,为各类高级系统(如MES、调度系统、库位管理、立库等)提供高效便捷的语音交互体验。
L…

(LeetCode 每日一题) 3442. 奇偶频次间的最大差值 I (哈希、字符串)
题目:3442. 奇偶频次间的最大差值 I 思路 :哈希,时间复杂度0(n)。 用哈希表来记录每个字符串中字符的分布情况,哈希表这里用数组即可实现。
C版本:
class Solution {
public:int maxDifference(string s) {int a[26]…

【大模型RAG】拍照搜题技术架构速览:三层管道、两级检索、兜底大模型
摘要
拍照搜题系统采用“三层管道(多模态 OCR → 语义检索 → 答案渲染)、两级检索(倒排 BM25 向量 HNSW)并以大语言模型兜底”的整体框架: 多模态 OCR 层 将题目图片经过超分、去噪、倾斜校正后,分别用…
【Axure高保真原型】引导弹窗
今天和大家中分享引导弹窗的原型模板,载入页面后,会显示引导弹窗,适用于引导用户使用页面,点击完成后,会显示下一个引导弹窗,直至最后一个引导弹窗完成后进入首页。具体效果可以点击下方视频观看或打开下方…
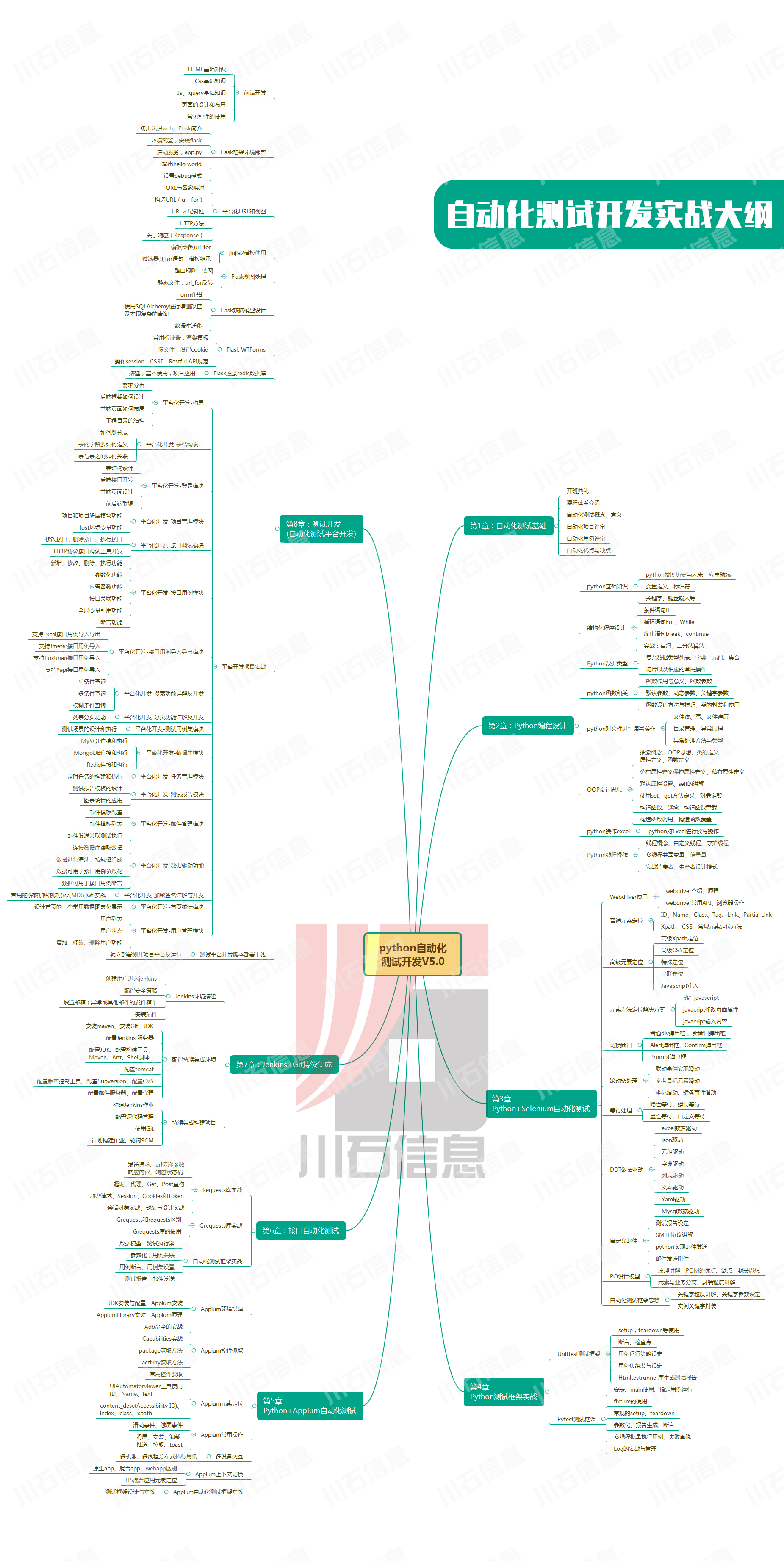
接口测试中缓存处理策略
在接口测试中,缓存处理策略是一个关键环节,直接影响测试结果的准确性和可靠性。合理的缓存处理策略能够确保测试环境的一致性,避免因缓存数据导致的测试偏差。以下是接口测试中常见的缓存处理策略及其详细说明:
一、缓存处理的核…
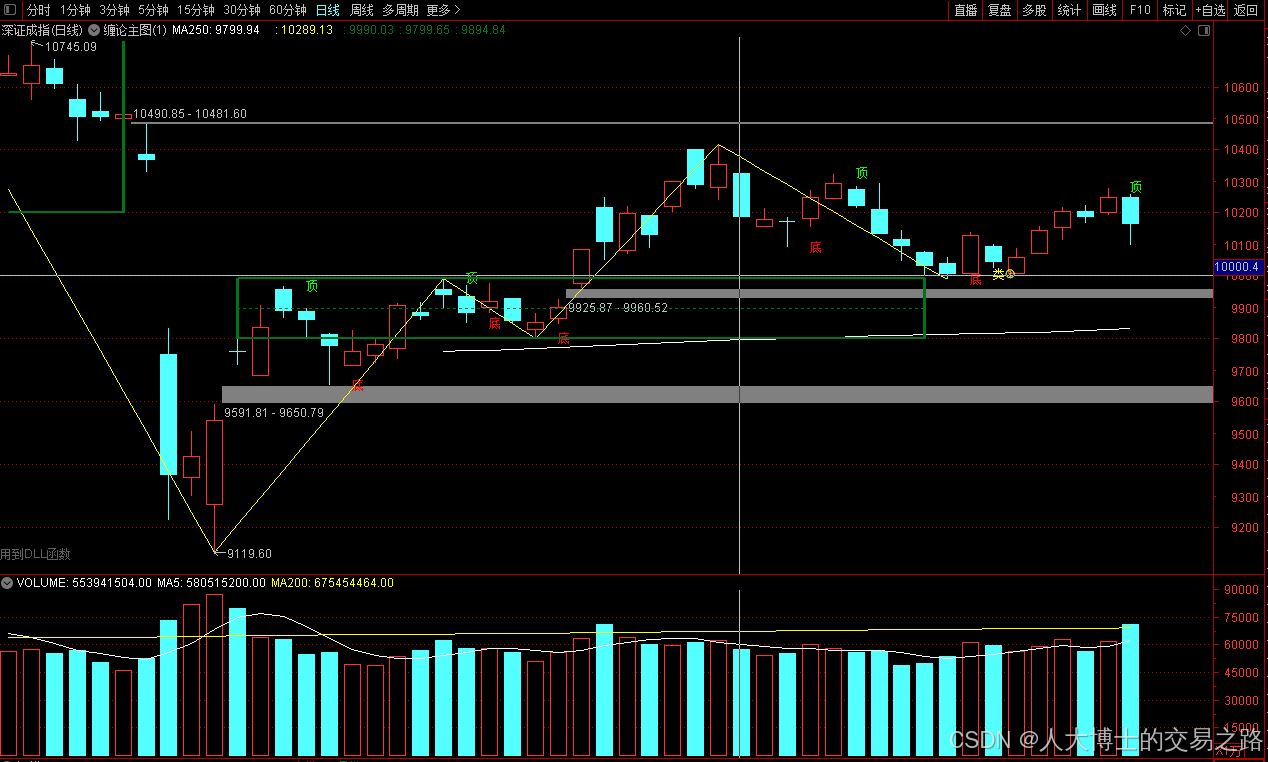
龙虎榜——20250610
上证指数放量收阴线,个股多数下跌,盘中受消息影响大幅波动。 深证指数放量收阴线形成顶分型,指数短线有调整的需求,大概需要一两天。 2025年6月10日龙虎榜行业方向分析 1. 金融科技
代表标的:御银股份、雄帝科技
驱动…
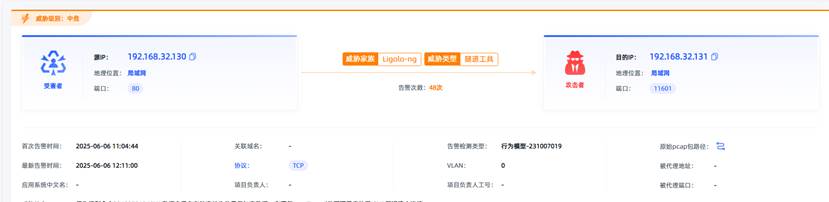
观成科技:隐蔽隧道工具Ligolo-ng加密流量分析
1.工具介绍
Ligolo-ng是一款由go编写的高效隧道工具,该工具基于TUN接口实现其功能,利用反向TCP/TLS连接建立一条隐蔽的通信信道,支持使用Let’s Encrypt自动生成证书。Ligolo-ng的通信隐蔽性体现在其支持多种连接方式,适应复杂网…

铭豹扩展坞 USB转网口 突然无法识别解决方法
当 USB 转网口扩展坞在一台笔记本上无法识别,但在其他电脑上正常工作时,问题通常出在笔记本自身或其与扩展坞的兼容性上。以下是系统化的定位思路和排查步骤,帮助你快速找到故障原因:
背景:
一个M-pard(铭豹)扩展坞的网卡突然无法识别了,扩展出来的三个USB接口正常。…

未来机器人的大脑:如何用神经网络模拟器实现更智能的决策?
编辑:陈萍萍的公主一点人工一点智能 未来机器人的大脑:如何用神经网络模拟器实现更智能的决策?RWM通过双自回归机制有效解决了复合误差、部分可观测性和随机动力学等关键挑战,在不依赖领域特定归纳偏见的条件下实现了卓越的预测准…

Linux应用开发之网络套接字编程(实例篇)
服务端与客户端单连接
服务端代码
#include <sys/socket.h>
#include <sys/types.h>
#include <netinet/in.h>
#include <stdio.h>
#include <stdlib.h>
#include <string.h>
#include <arpa/inet.h>
#include <pthread.h>
…

华为云AI开发平台ModelArts
华为云ModelArts:重塑AI开发流程的“智能引擎”与“创新加速器”!
在人工智能浪潮席卷全球的2025年,企业拥抱AI的意愿空前高涨,但技术门槛高、流程复杂、资源投入巨大的现实,却让许多创新构想止步于实验室。数据科学家…

深度学习在微纳光子学中的应用
深度学习在微纳光子学中的主要应用方向
深度学习与微纳光子学的结合主要集中在以下几个方向:
逆向设计 通过神经网络快速预测微纳结构的光学响应,替代传统耗时的数值模拟方法。例如设计超表面、光子晶体等结构。
特征提取与优化 从复杂的光学数据中自…